Download
1 / 1
10 likes | 375 Views
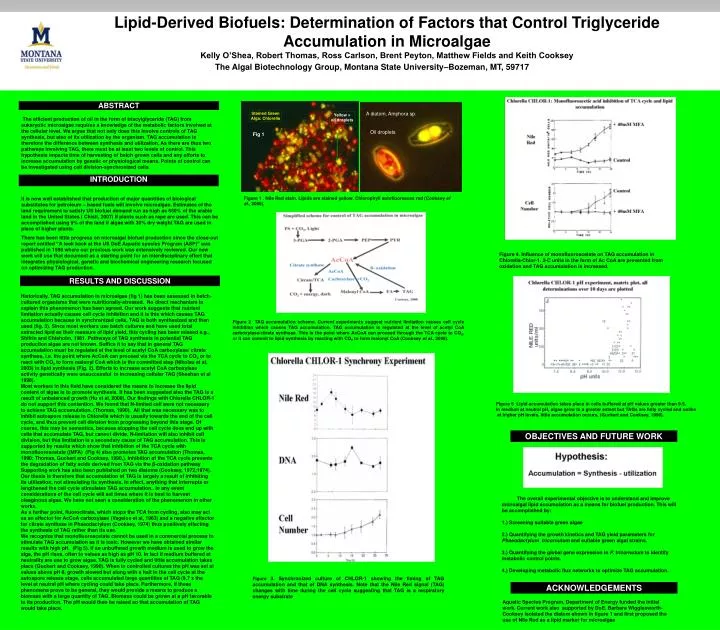
Stained Green Alga: Chlorella. Yellow = oil droplets. Lipid-Derived Biofuels: Determination of Factors that Control Triglyceride Accumulation in Microalgae Kelly O’Shea, Robert Thomas, Ross Carlson, Brent Peyton, Matthew Fields and Keith Cooksey.

E N D
Stained Green Alga: Chlorella Yellow = oil droplets Lipid-Derived Biofuels: Determination of Factors that Control Triglyceride Accumulation in Microalgae Kelly O’Shea, Robert Thomas, Ross Carlson, Brent Peyton, Matthew Fields and Keith Cooksey The Algal Biotechnology Group, Montana State University–Bozeman, MT, 59717 ABSTRACT A diatom, Amphora sp. The efficient production of oil in the form of triacylglyceride (TAG) from eukaryotic microalgae requires a knowledge of the metabolic factors involved at the cellular level. We argue that not only does this involve controls of TAG synthesis, but also of its utilization by the organism. TAG accumulation is therefore the difference between synthesis and utilization. As there are thus two pathways involving TAG, there must be at least two levels of control. This hypothesis impacts time of harvesting of batch grown cells and any efforts to increase accumulation by genetic or physiological means. Points of control can be investigated using cell division-synchronized cells. Oil droplets Fig 1 INTRODUCTION Figure 1 . Nile Red stain. Lipids are stained yellow. Chlorophyll autofluoresces red (Cooksey et al., 2008). It is now well established that production of major quantities of biological substitutes for petroleum – based fuels will involve microalgae. Estimates of the land requirement to satisfy US biofuel demand run as high as 650% of the arable land in the United States ( Chisti, 2007) if plants such as rape are used. This can be accomplished using 9% of the land if algae with 30% dry weight TAG are used in place of higher plants. There has been little progress on microalgal biofuel production since the close-out report entitled “A look back at the US DoE Aquatic species Program (ASP)” was published in 1998 where our previous work was extensively reviewed. Our new work will use that document as a starting point for an interdisciplinary effort that integrates physiological, genetic and biochemical engineering research focused on optimizing TAG production. Figure 4. Influence of monofluoroacetate on TAG accumulation in Chlorella-Chlor-1. 2-C units in the form of Ac CoA are prevented from oxidation and TAG accumulation is increased. RESULTS AND DISCUSSION Historically, TAG accumulation in microalgae (fig 1) has been assessed in batch-cultured organisms that were nutritionally-stressed. No direct mechanism to explain this phenomenon has been agreed. Our work suggests that nutrient limitation actually causes cell cycle inhibition and it is this which causes TAG accumulation because in synchronized cells, TAG is both synthesized and then used (fig. 3). Since most workers use batch cultures and have used total extracted lipid as their measure of lipid yield, this cycling has been missed e.g., Shifrin and Chisholm, 1981. Pathways of TAG synthesis in potential TAG production algae are not known. Suffice it to say that in general TAG accumulation must be regulated at the level of acetyl CoA carboxylase/ citrate synthase, i.e. the point where AcCoA can proceed via the TCA cycle to CO2 or to react with CO2 to form malonyl CoA which is the committed step (Nikolau et al, 2003) in lipid synthesis (Fig. 2). Efforts to increase acetyl CoA carboxylase activity genetically were unsuccessful in increasing cellular TAG (Sheehan et al 1998). Most workers in this field have considered the means to increase the lipid content of algae is to promote synthesis. It has been suggested also the TAG is a result of unbalanced growth (Hu et al, 2008). Our findings with Chlorella CHLOR-1 do not support this contention. We found that N-limited cell were not necessary to achieve TAG accumulation. (Thomas, 1990). All that was necessary was to inhibit autospore release in Chlorella which is usually towards the end of the cell cycle, and thus prevent cell division from progressing beyond this stage. Of course, this may be semantics, because stopping the cell cycle does end up with cells that accumulate TAG, but cannot divide. N-limitation will also inhibit cell division, but this limitation is a secondary cause of TAG accumulation. This is supported by results which show that inhibition of the TCA cycle with monofluoroacetate ((MFA) (Fig 4) also promotes TAG accumulation (Thomas, 1990; Thomas, Guckert and Cooksey, 1990,). Inhibition of the TCA cycle prevents the degradation of fatty acids derived from TAG via the β-oxidation pathway Supporting work has also been published on two diatoms (Cooksey, 1972;1974). Our thesis is therefore that accumulation of TAG is largely a result of inhibiting its utilization, not stimulating its synthesis. In effect, anything that interrupts or lengthened the cell cycle stimulates TAG accumulation.. In any event considerations of the cell cycle will set times where it is best to harvest oleaginous algae. We have not seen a consideration of the phenomenon in other works. As a further point, fluorocitrate, which stops the TCA from cycling, also may act as an effector for AcCoA carboxylase (Vagelos et al, 1963) and a negative effector for citrate synthase in Phaeodactylum (Cooksey, 1974) thus positively effecting the synthesis of TAG rather than its use.` We recognize that monofluoroacetate cannot be used in a commercial process to stimulate TAG accumulation as it is toxic. However we have obtained similar results with high pH. (Fig 5). If an unbuffered growth medium is used to grow the alga, the pH rises, often to values as high as pH 10. In fact if medium buffered at neutrality are use to grow algae, TAG is fully cycled and little accumulation takes place (Guckert and Cooksey, 1990). When in controlled cultures the pH was set at values above pH 8, growth slowed but along with a halt in the cell cycle at the autospore release stage, cells accumulated large quantities of TAG (9.7 x the level at neutral pH where cycling could take place. Furthermore, if these phenomena prove to be general, they would provide a means to produce a biomass with a large quantity of TAG. Biomass could be grown at a pH favorable to its production. The pH would then be raised so that accumulation of TAG would take place. Figure 2 TAG accumulation scheme. Current experiments suggest nutrient limitation causes cell cycle inhibition which causes TAG accumulation. TAG accumulation is regulated at the level of acetyl CoA carboxylase/citrate synthase. This is the point where AcCoA can proceed through the TCA cycle to CO2, or it can commit to lipid synthesis by reacting with CO2 to form malonyl CoA (Cooksey et al., 2008). Figure 5 Lipid accumulation takes place in cells buffered at pH values greater than 9.5. In medium at neutral pH, algae grow to a greater extent but TAGs are fully cycled and unlike at higher pH levels, little accumulation occurs, (Guckert and Cooksey, 1990). OBJECTIVES AND FUTURE WORK The overall experimental objective is to understand and improve microalgal lipid accumulation as a means for biofuel production. This will be accomplished by: 1.) Screening suitable green algae 2.) Quantifying the growth kinetics and TAG yield parameters for Phaeodactylum tricornutum and suitable green algal strains. 3.) Quantifying the global gene expression in P. tricornutum to identify metabolic control points. 4.) Developing metabolic flux networks to optimize TAG accumulation. Figure 3. Synchronized culture of CHLOR-1 showing the timing of TAG accumulation and that of DNA synthesis. Note that the Nile Red signal (TAG) changes with time during the cell cycle suggesting that TAG is a respiratory energy substrate ACKNOWLEDGEMENTS Aquatic Species Program, Department of Energy funded the initial work. Current work also supported by DoE. Barbara Wigglesworth-Cooksey isolated the diatom shown in figure 1 and first proposed the use of Nile Red as a lipid marker for microalgae