Download

1 / 67
670 likes | 685 Views
Discover the biochemical basis of muscle movement and explore molecular motors like Dynein, Kinesin, and helicases. Learn about the chemistry of muscle contraction, sliding filament model, and the processivity of motor proteins.

E N D
Motors II;Muscle Contraction Andy HowardIntroductory Biochemistry1 December 2014 Motors II; Muscle Biochemistry
Chemistry of muscle contraction • The most impressive movement phenomenon in mesoscopic organisms is muscle movement. It does have a biochemical basis, which we’ll explore today. • … But first we’ll finish our conversation about other molecular motors Motors II; Muscle Biochemistry
Other motors Dynein, Kinesin Helicases Processivity Flagella revisited Skeletal muscle Thin filaments: actin, tropomyosin, troponin Thick filaments: myosin Sliding filament model Dystrophin and cytoskeletal structure Coupling Myosin & kinesin Calcium channels and troponin C, I Smooth muscle What we’ll discuss Motors II; Muscle Biochemistry
Movement of organelles and vacuoles • Can be fast:2-5 µm s-1 • Hard to study • 1985: Kinesin isolated • 1987: Cytosolic dynein found Motors II; Muscle Biochemistry
Dynein movement Motors II; Muscle Biochemistry
Cytosolic dynein • Mostly moves organelles & vesicles from (+) to (-), so it moves things toward the center of the cell • Heavy chain ~ 400kDa, plus smaller peptides (53-74 kDa) • Microtubule-activated ATPase activity Motors II; Muscle Biochemistry
Kinesin • Mostly moves organelles from (-) to (+) • That has the effect of moving things outward • 360 kDa: 110 kDa heavy chains, also 65-70 kDa subunits (2 + 2?) • Head domain of heavy chain (38 kDa) binds ATP and microtubule: cooperative interactions between pairs of head domains in kinesin, causing conformational changes in a single tubulin subunit • 8 nm movements along long axis of microtubule Motors II; Muscle Biochemistry
Kinesin motion depicted • Rolling movement involving two head domains at a time • Fig. 16.8(b) Motors II; Muscle Biochemistry
Hand-over-hand kinesin model • Two head groups begin in contact • After ATP hydrolysis hindmost head passes forward head • ATP binds to new leading head • Pi dissociates from trailing head Motors II; Muscle Biochemistry
DNA helicases • To replicate DNA we need to separate the strands • Efficient only if the helicase can travel along the duplex quickly • This kind of movement is called processive • E.coli BCD helicase can unwind 33kbp before it falls off • If we want to replicate DNA rapidly, we need processivity Motors II; Muscle Biochemistry
Achieving processivity • Some helicases form rings that encircle 1 or both strands of the duplex • Others, like rep helicase, are homodimeric; move hand-over-hand along the DNA, like kinesin Motors II; Muscle Biochemistry
Negative cooperativity • Rep is monomeric without DNA • Each monomer can bind either ss or dsDNA • BUT after one monomer binds DNA, the second subunit’s affinity drops 104-fold! Motors II; Muscle Biochemistry
Bacterial flagella • E.coli flagellum is 10 µm in length,15 nm in diameter • ~6 filaments on surface of cell rotate counter-clockwise: that makes them bundle together and propel the cell through medium • Enabled by rotation of motor protein complexes in plasma membrane Motors II; Muscle Biochemistry
Motor structure • ≥ 2 rings, ~25nm diameter (M & S) • Rod attaches those to the helical filament • Rings surrounded by array of membrane proteins • This one is driven by a proton gradient, not by ATP hydrolysis:[H+]out > [H+]in, so protons want to move in • If we let protons in, we can use the thermodynamic energy to drive movement • Requires 800-1200 protons per full rotation!… that’s equivalent to ~ 250 ATP’s Motors II; Muscle Biochemistry
The shuttle • MotA & MotB form shuttling device • Proton movement drives rotation of flagellar motor • Fig. 16.26 Motors II; Muscle Biochemistry
iClicker question 1 1. Compare the pH inside a typical bacterial cell to the pH outside. • (a) pHin < pHout • (b) pHin> pHout • (c) pHin = pHout • (d) We don’t have enough information to answer this question. Motors II; Muscle Biochemistry
Berg’s model • motB has protonexchanging sites • motA has half-channels—one half facing toward the inside of the cell, one facing out • When a motB site is protonated, the outside edges of motA can’t move past it • Center of motA can’t move past site when it’s empty • Those constraints cause coupling between proton translocation and rotation Motors II; Muscle Biochemistry
Coupling described • Proton enters outside of motA and binds to an exchange site on motB • motA is linked to cell wall, so when it rotates, it puts the inside channel over the proton • Proton moves through inside channel into cell; then another proton travels up the outside channel to bind to the next exchange site • That pulls the complex to the left, leading to counterclockwise rotation of disc, rod, & helical filament Motors II; Muscle Biochemistry
Coupling depicted • Fig. 16.27 Motors II; Muscle Biochemistry
What if it got reversed? • If outside became alkaline, the flagellar filaments would rotate clockwise • That doesn’t work as well because it loosens the microtubule Motors II; Muscle Biochemistry
Quantitation • M ring has about 100 motB exchange sites • 800-1200 protons for a full rotation of the filament • That enables ~ 100 rotations/sec Motors II; Muscle Biochemistry
Muscle contraction • This is an obvious case of an energy-dependent biological motion system • Involves an interaction called the sliding filament model, in which myosin molecules slide past actin molecules • Many other proteins and structural components involved Motors II; Muscle Biochemistry
Essential Question • How can biological macromolecules, carrying out conformational changes on the microscopic, molecular level, achieve these feats of movement that span the molecular and macroscopic worlds? • We’ll look at the specifics of muscle contraction, which is an excellent example of this phenomenon • Note that Tom Irving, on our faculty, is a world-recognized expert on muscle physiology Prof. Thomas C. Irving Motors II; Muscle Biochemistry
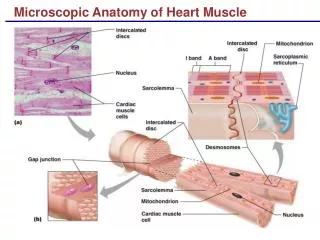
Skeletal Muscle Cell G&G fig. 16.1 • T-tubules enable the sarcolemmal membrane to contact the ends of the myofibril Motors II; Muscle Biochemistry
What are t-tubules (transverse tubules) and the sarcoplasmic reticulum for? • The morphology is all geared to Ca2+ release and uptake! • Nerve impulses reaching the muscle produce an "action potential" that spreads over the sarcolemmal membrane and into the fiber along the t-tubule network Motors II; Muscle Biochemistry
t-tubules and SR, continued • The signal is passed across the triad junction and induces release of Ca2+ ions from the SR • Ca2+ ions bind to sites on the fibers and induce contraction; relaxation involves pumping the Ca2+ back into the SR Motors II; Muscle Biochemistry
Molecular mechanism of contraction Be able to explain the EM in Figure 16.2in terms of thin and thick filaments • Thin filaments are composed of actin polymers • F-actin helix is composed of G-actin monomers • F-actin helix has a pitch of 72 nm • But repeat distance is 36 nm • Actin filaments are decorated with tropomyosin heterodimers and troponin complexes • Troponin complex consists of: troponin T (TnT), troponin I (TnI), and troponin C (TnC) Motors II; Muscle Biochemistry
Myo- fibrils • Hexagonal arrays shown Motors II; Muscle Biochemistry
Actin monomer • One domain on each side(fig. 16.3) Motors II; Muscle Biochemistry
Actin helices • Pitch = 72nm • Repeat = 36 nm • Cf. Fig.16.4 Motors II; Muscle Biochemistry
Thin filament • Tropomyosin coiled coil winds around the actin helix • Each TM dimer interacts with 7 actin monomers • Troponin T binds to TM at head-to-tail junction Motors II; Muscle Biochemistry
Composition & Structure of Thick Filaments Myosin - 2 heavy chains, 4 light chains • Heavy chains - 230 kD each • Light chains - 2 pairs of different 20 kD chains • The "heads" of heavy chains have ATPase activity and hydrolysis here drives contraction • Light chains are homologous to calmodulin and also to TnC • See structure of heads in the G&G figure Motors II; Muscle Biochemistry
Myosin • Cartoon • EM • S1 myosin head structure • Cf. Fig. 16.5(b) Motors II; Muscle Biochemistry
Repeating Structural Elements Are the Secret of Myosin’s Coiled Coils • 7-residue, 28-residue and 196-residue repeats are responsible for the organization of thick filaments • Residues 1 and 4 (a and d) of the seven-residue repeat are hydrophobic; residues 2,3 and 6 (b, c and f) are ionic • This repeating pattern favors formation of coiled coil of tails. (With 3.6 - NOT 3.5 - residues per turn, α-helices will coil!) Motors II; Muscle Biochemistry
iClicker question 2 • 2. The lowest-level repeat pattern involves 7 residues; the highest is 196 residues. How many lowest-level repeats are contained in one highest-level repeat? • (a) 196 • (b) 28 • (c) 14 • (d) 7 • (e) undeterminable from the data given. Motors II; Muscle Biochemistry
Axial view Myosin tail: 2-stranded -helical coiled coil Motors II; Muscle Biochemistry
More Myosin Repeats! • 28-residue repeat (4 x 7) consists of distinct patterns of alternating side-chain charge (+ vs -), and these regions pack with regions of opposite charge on adjacent myosins to stabilize the filament • 196-residue repeat (7 x 28) pattern also contributes to packing and stability of filaments Motors II; Muscle Biochemistry
Myosin packing • Adjoining molecules offset by ~ 14 nm • Corresponds to 98 residues of coiled coil Motors II; Muscle Biochemistry
Associated proteins of Muscle • -Actinin, a protein that contains several repeat units, forms dimers and contains actin-binding regions, and is analogous in some ways to dystrophin • Dystrophin is the protein product of the first gene to be associated with muscular dystrophy - actually Duchennes MD • See the boxes on this subject in text Motors II; Muscle Biochemistry
Dystrophin Dystrophin is part of a large complex of glycoproteins that bridges the inner cytoskeleton (actin filaments) and the extracellular matrix (via a protein called laminin) • Two subcomplexes: dystroglycan and sarcoglycan • Defects in these proteins have now been linked to other forms of muscular dystrophy Nick Menhart:BCPS faculty member specializing in dystrophin research Motors II; Muscle Biochemistry
Dystrophin, actinin,spectrin • Characteristic 3-helix regions • Cf. box on pp. 522-523 Motors II; Muscle Biochemistry
Spectrin-repeat structure • These characteristic 3-helix elements are found in actinin, spectrin, dystrophin Human dystrophin: first spectrin repeat27 kDa dimer;monomer shownPDB 3UUN, 2.3Å Motors II; Muscle Biochemistry
Model for complex • Actin-dystrophin-glycoprotein complex • Dystrophin forms tetramers of antiparallel monomers Motors II; Muscle Biochemistry
The Dystrophin Complex Links to disease • -Dystroglycan - extracellular, binds to merosin (a component of laminin) - mutation in merosin linked to severe congenital muscular dystrophy • -Dystroglycan - transmembrane protein that binds dystrophin inside • Sarcoglycan complex - , , - all transmembrane - defects linked to limb-girdle MD and autosomal recessive MD Motors II; Muscle Biochemistry
Hugh Huxley The Sliding Filament Model Many contributors! • Hugh Huxley and Jean Hanson • Andrew Huxley and Ralph Niedergerke • Albert Szent-Györgyi showed that actin and myosin associate (actomyosin complex) • Sarcomeres decrease length during contraction (see Figure 16.19) • Szent-Györgyi also showed that ATP causes the actomyosin complex to dissociate Albert Szent-Györgyi Motors II; Muscle Biochemistry
Sliding filaments • Decrease in sarcomere length happens because of decreases in width of I band and H zone • No change in width of A band • Thin & thick filaments are sliding past one another Motors II; Muscle Biochemistry
The Contraction Cycle Study Figure 16.9! • Cross-bridge formation is followed by power stroke with ADP and Pi release • ATP binding causes dissociation of myosin heads and reorientation of myosin head • Details of the conformational change in the myosin heads are coming to light! • Evidence now exists for a movement of at least 35 Å in the conformation change between the ADP-bound state and ADP-free state Motors II; Muscle Biochemistry
Mechanism • Fig. 16.9 Motors II; Muscle Biochemistry
Actin-myosin interaction • Ribbon- and space-filling representations Ivan Rayment Hazel Holden Motors II; Muscle Biochemistry
Similarities in Motor Proteins • Initial events of myosin and kinesin action are similar • But the conformational changes that induce movement are different in myosins, kinesins, and dyneins Motors II; Muscle Biochemistry