Download
1 / 55
550 likes | 756 Views
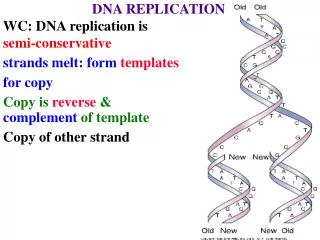
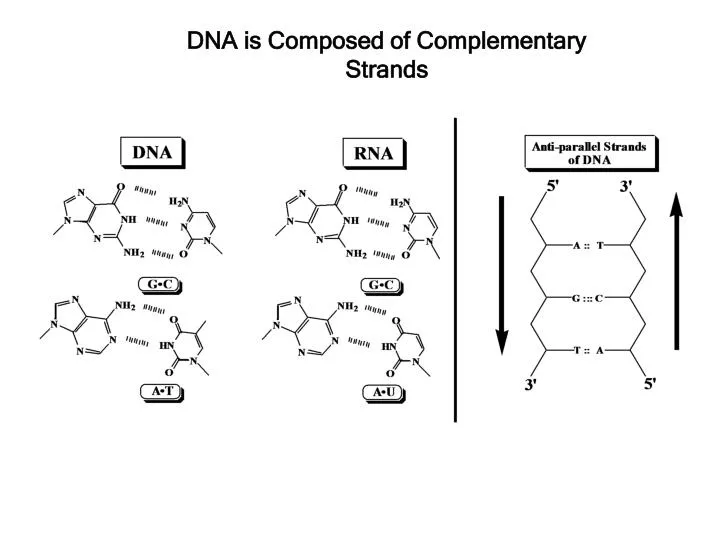
DNA is Composed of Complementary Strands. Base Pairing is Determined by Hydrogen Bonding. same size. Forces stabilizing DNA double helix. Hydrogen bonding (2-3 kcal/mol per base pair) Stacking (hydrophobic) interactions (4-15 kcal/mol per base pair) 3. Electrostatic forces. B-DNA.

E N D
Forces stabilizing DNA double helix • Hydrogen bonding (2-3 kcal/mol per base pair) • Stacking (hydrophobic) interactions • (4-15 kcal/mol per base pair) • 3. Electrostatic forces.
B-DNA • •Sugars are in the 2’ endo conformation. • •Bases are the anti conformation. • •Bases have a helical twist of 36º • (10.4 bases per helix turn) • Helical pitch = 34 A 23.7 A right handed helix • helical axis passes through • base pairs 7.0 A • planes of bases are nearly • perpendicular to the helix axis. • 3.4 A rise between base pairs Wide and deep Narrow and deep
DNA can deviate from Ideal Watson-Crick structure • Helical twist ranges from 28 to 42° • Propeller twisting 10 to 20° • Base pair roll
N NH 2 H N O 2 N N HN C-1’ N N NH O 2 C-1’ Major and minor groove of the double helix O N NH N N N N C-1’ O C-1’ Wide and deep Narrow and deep
Major groove and Minor groove of DNA N NH O 2 N H N O 2 N NH N N N HN C-1’ N N N N C-1’ NH O O 2 C-1’ Hypothetical situation: the two grooves would have similar size if dR residues were attached at 180° to each other To deoxyribose-C1’ C1’ -To deoxyribose C-1’
B-type duplex is not possible for RNA steric “clash”
A-form helix:dehydrated DNA; RNA-DNA hybrids • •Sugars are in the 3’ endo conformation. • •Bases are the anti conformation. • •11 bases per helix turn • Helical pitch = 25.3 A Right handed helix • planes of bases are tilted • 20 ° relative the helix axis. • 2.3 A rise between base pairs 25.5 A Top View
The sugar puckering in A-DNA is 3’-endo 5.9 A 7.0 A
Living Figure – A-DNA http://bcs.whfreeman.com/biochem5
A-DNA has a shallow minor groove and a deep major groove • • Helix axis A-DNA B-DNA
Z-form double helix:polynucleotides of alternating purines and pyrimidines (GCGCGCGC) at high salt • • Backbone zig-zags because sugar puckers alternate between 2’ endo pyrimidines and 3’ endo (purines) • • Bases alternate between anti (pyrimidines) and syn conformation (purines). • •12 bases per helix turn • Helical pitch = 45.6 A Left handed helix • planes of the bases are • tilted 9° relative the helix • axis. • 3.8 A rise between base pairs 18.4 A • Flat major groove • Narrow and deep minor groove
Sugar and base conformations in Z-DNA alternate: 5’-GCGCGCGCGCGCG 3’-CGCGCGCGCGCGC C:sugar is 2’-endo, base is anti G: sugar is 3’-endo, base is syn
Living Figure – Z-DNA http://bcs.whfreeman.com/biochem5
Biological relevance of the minor types of DNA secondary structure • Although the majority of chromosomal DNA is in B-form, • some regions assume A- or Z-like structure • Runs of multiple Gs are A-like • The upstream sequences of some genes contain • 5-methylcytosine = Z-like duplex • Structural variations play a role in DNA-protein interactions • RNA-DNA hybrids and ds RNA have an A-type structure
Hydrogen bond donors and acceptors in DNA grooves facilitate its recognition by proteins H N H N 2 2 The edges of base pairs displayed to DNA major and minor groove contain potential H-bond donors and acceptors: O N n h o h n= Nitrogen hydrogen bond acceptor o= Oxygen hydrogen bond acceptor h= Amino hydrogen bond donor
Hydrogen bond donors and acceptors on each edge of a base pair
Structural characteristics of DNA facilitating DNA-Protein Recogtnition • Major and major groove of DNA contain sequence- • dependent patterns of H-bond donors and acceptors. • Sequence-dependent duplex structure (A, B, Z, bent • DNA). • Hydrophobic interactions via intercalation. • Ionic interactions with phosphates.
Leucine zipper proteins bind DNA major groove Groove binding drugs and proteins 5’-ATT-3’ Others: netropsin, distamycin, Hoechst 33258
Triple helix and Antigene approach Hoogsteen base pairing = parallel Reversed Hoogsteen = antiparallel
Biophysical properties of DNA • Facile denaturation (melting) and re-association of the duplex • are important for DNA’s biological functions. • In the laboratory, melting can be induced by heating. Single strands T° duplex • Hybridization techniques are based on the affinity of complementary • DNA strands for each other. • Duplex stability is affected by DNA length, % GC base pairs, ionic strength, the presence of organic solvents, pH • Negative charge – can be separated by gel electrophoresis
Separation of DNA fragments by gel electrophoresis Polyacrylamide gel: • DNA strands are negatively charged – • migrate towards the anode • Migration time ~ ln (number of base • pairs)
DNA Topology DNA has to be coiled to fit inside the cell DNA polymers must be folded to fit into the cell or nucleus (tertiary structure).
Connect the ends to make a circular DNA: Tw = 260/10.4 = 25
An electron micrograph of negatively supercoiled and relaxed DNA Stryer Fig. 27.20
Organization of chromosomal DNA • Chromosomal DNA is organized in loops (no free ends) • It is negatively supercoiled: 1 (-) supercoil per 200 nucleotides 145 bp duplex Histone octamer (H2A, H2B, H3, H4)2 H1 is bound to the linker region
Enzymes that control DNA supercoiling: DNA Topoisomerases Change the linking number (Lk) of DNA duplex by concerted breakage and re-joining DNA strands Topoisomerase enzymes Topoisomerases I Relax DNA supercoiling by increments of 1 (cleave one strand) Topoisomerases II Change DNA supercoiling by the increments of 2 (break both strands) Usually introduce negative supercoiling
Human DNA Topoisomerase I: DNA: side view 20Å Stryer Fig. 27.21
Mechanism of DNA Topoisomerases I 723 OH P-Topo Wr = 1
Drugs that inhibit DNA Topoisomerase I • • Camptothecin, topotecan and analogs • •Antitumor activity correlates with interference with topoisomerase activity • • Stabilizes topoisomerase I-DNA intermediate, preventing DNA strand re-ligation • Used in treatment of colorectal, ovarian, and small cell lung tumors
Enzymes that control DNA supercoiling: DNA Topoisomerases Change the linking number (Lk) of DNA duplex by concerted breakage and re-joining DNA strands Topoisomerase enzymes Topoisomerases I Relax DNA supercoiling by increments of 1 (cleave one strand) Topoisomerases II Change DNA supercoiling by the increments of 2 (break both strands) Usually introduce negative supercoiling
Topoisomerases II • Most of Topoisomerases II introduce negative supercoils (e.g. E. coli DNA Gyrase) • Require energy (ATP) • Each round introduces two supercoils ( Wr = - 2) • Necessary for DNA synthesis • Form a covalent DNA-protein complex similar to Topoisomerases I
Yeast DNA Topoisomerase II Stryer Fig. 27.23
Topoisomerase II - mechanism Stryer Fig. 27.24
Drugs that inhibit bacterial Topoisomerase II (DNA gyrase) Interfere with breakage and rejoining DNA ends: Inhibit ATP binding:
Enzymes that cut DNA: exonucleases Phosphate group Nucleobase 2’-deoxyribose HO 5’ A OH 3’ 5’ 3’ 5’ + dNMPs • Degrade DNA in a stepwise manner by removing deoxynucleotides in 5’ 3’ (A) or 3’ 5’ direction (B) • Require a free OH • Most exonucleases are active on both single- and double-stranded DNA • Used for degrading foreign DNA and in proofreading during DNA synthesis B HO H 3’
DNA Endonucleases • Cleave internal phosphodiester bonds resulting in 3’-OH and 5’-phosphate ends 5’ 3’-OH 5’-P 5’-P 3’-OH • some endonucleases cleave randomly (DNase I, II) • Type II Restriction endonucleases are highly sequence specific EcoRI recognition site: Palindromic site (inverted repeat) • RE are found in bacteria where they are used for protection against foreign DNA
Recognition sequences of some common restriction endonucleases
Applications of Restriction Endonucleases in Molecular Biology • DNA fingerprinting (restriction fragment length polymorphism). • 2. Molecular cloning (isolation and amplification of genes).
Restriction fragment length polymorphisms are used to compare DNA from different sources