Download
1 / 18
180 likes | 320 Views
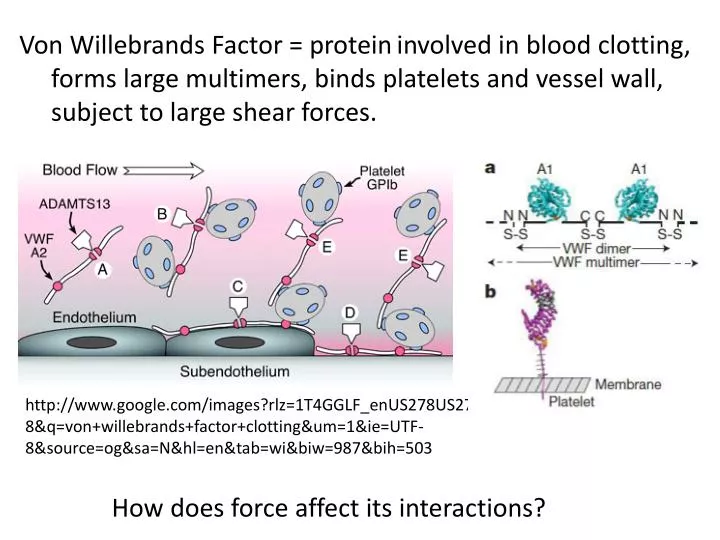
Von Willebrands Factor = protein involved in blood clotting, forms large multimers, binds platelets and vessel wall, subject to large shear forces. .

E N D
Von Willebrands Factor = proteininvolved in blood clotting, forms large multimers, binds platelets and vessel wall, subject to large shear forces. http://www.google.com/images?rlz=1T4GGLF_enUS278US278&q=von+willebrands+factor+clotting&um=1&ie=UTF-8&source=og&sa=N&hl=en&tab=wi&biw=987&bih=503 How does force affect its interactions?
Novel experimental set-up to study effect of force on interaction between A1 and GPIba
Experimental readout of force at which domains separate Note they are watching individual molecules they can repeatedly test the same molecule unbinding and rebinding forces are stochastic
Some technical details How long is linker? 43 amino acids, ~16nm fully extended Has to be long enough to see rupture Why use DNA “handles”? To keep proteins off surfaces, separate laser trapped bead from rest of assembly, apply force at single point How to attach proteins to DNA? via S-S bonds with extra unique cys in protein and SH in synthetic DNA
How does the laser trap work? Watch videos at http://tweezerslab.unipr.it or http://www.youtube.com/watch?v=BL9gmMzpRr4&feature=PlayList&p=DB423384C9BB437D&index=1 for more detailed explanation of trap set-up used here Idea: refraction changes direction (momentum) of light Dp Since momentum is conserved, bead’s momentum changes by – Dp F = -dDp/dt, mainly directed toward most intense part of beam
Optics – counterpropagating laser traps beam position force Infer bead position from force, knowing stiffness, x=F/k
Relation between force and Dextension Dx = length of “open” A1-linker-GPIba + DNA - length of A1-GPIba complex + DNA DNA length should be constant at given F If A1 and GPIba domains don’t stretch, Dx due to extension of flexible aa linker and rot. of A1, GPIba Bin data by force, show mean and variance of Dx at rupture or rebinding Results c/w expected stretch in linker w/force, hence, c/w single tether
How does loading rate, F(t), affect dn of unbinding forces? Rate of escape at F=0 ~e-DE/kT For constant pulling force F, energy barrier decreases by FDx, so rate ~e-(DE-FDx)/kT => av. lifetime <t(F)> = <t(0)> e-F(Dx/kT) = e-Fs/kT E DE If F changes with time, F(t), dn of unbinding forces changes, and need careful Dx pulling direction x analysis to determine relationship between probability of unbinding at force f for a given F(t) and the average lifetime at constant force, t(F), which should depend exp’lly on F
Dudko-Hummer-Szabo provide this analysis (PNAS 105:15755 (2008)) Relation between p(f) given F(t) and t(F) is: If data grouped into force bins of size DF; hi = p(rupture in ith force bin); in kth bin, F = F0+(k-1/2)DF = evaluated at F=F0+(k-1/2)DF
Kim et al observe p(f) at different loading rates, find bimodal distributions at intermediate loading rates, then calculate <t(F)> for each peak according to D-H-S Obs. p(f) calculated <t(F)> k1(or2),off = t-1extrapolated to F=0
1/k2,off 2 Note ln t(F) ~ F for each peak as in simple theory Their interpretation: 2 different bound states, each with its own exp’l dependence on F (slope s/kT) and predicted zero force unbinding rate k1,off, k2,off 1/k1,off
Repeat expts at constant pulling F close to value at which bound/unbound states are equally likely, watch transitions back and forth B<->UB From these data calculate prob. of staying in bound state for time t
straight line => psurvival~ e-kt ; k = rate of unbinding exp’l survival prob. expected for process with single random event (like radioactive decay) Two exponentials at intermediate forces, c/w 2 bound states with different unbinding rates (that are fns of F)
Their model: 2 bound states interconvert at (force- dependent) rates k12 and k21estimated from data Note both bonds are weakened by force (s>0, “slip” rather than “catch”) but conversion to bond with lower koff and smaller s -> tighter binding at high shear (“flex” bond)
Ristocetin (antibiotic) and botrocetin (snake venom toxin) are drugs known to affect vWF They promoted conversion to state 2 and decreased response to force (s)
Main points Novel set-up to repeatedly measure interaction between 2 protein domains in single protein as function of force Nice “corroboration” of D-H-S theory that relates unbinding force distributions at different loading rates to lifetimes at constant force, with complication that… Their system had bimodal unbinding force distributions at some loading rates. This suggests protein complex exists in 2 different states, each state behaving as expected for simple model with ln(t(F)) = Fs/kT, but different sensitivities (s’s) for the 2 states
The 2 bound-state model is biologically interesting since it leads to a more graded response to force loading for an interaction naturally subject to comparable forces Will it turn out that many protein interactions have alter- native binding states? Single-molecule pulling experiments reveal complexities of protein interactions that would be hard to see in bulk assays Compared to AFM, newer pulling methods allow study in lower force regime (pN vs nN), and away from surfaces which can -> artifacts
Relevance to nanotechnology in other areas methods to detect forces ~0.1-10pN distance changes ~10nm ways to relate rupture-force distributions at different pulling rates to lifetimes at constant force ? interest from ME point of view – combining 2 “slip” bonds with different force sensitivities can mimic “catch” bond (increase in F -> sudden increase in bond strength)