Download
1 / 1
10 likes | 173 Views
NEURONAL NETWORK ACTIVITY IN THE LIMBIC CIRCUITRY IN TAU TRANSGENIC (TG4510) MICE Liam Scott, Tamás Kiss, Thomas T. Kawabe, Brian Harvey, Mihály Hajós # Neuroscience Research Unit, Pfizer Global Research & Development, Groton, Connecticut, U.S.A.

E N D
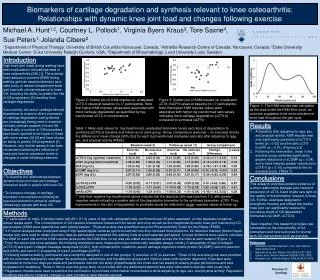
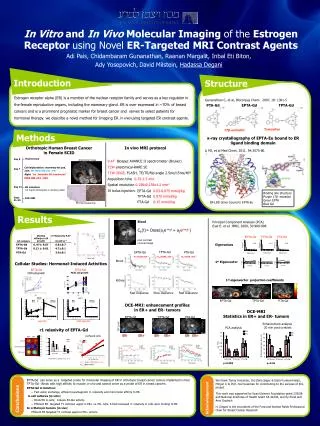
NEURONAL NETWORK ACTIVITY IN THE LIMBIC CIRCUITRY IN TAU TRANSGENIC (TG4510) MICE Liam Scott, Tamás Kiss, Thomas T. Kawabe, Brian Harvey, Mihály Hajós# Neuroscience Research Unit, Pfizer Global Research & Development, Groton, Connecticut, U.S.A. # Current affiliation: Translational Neuropharmacology, Section of Comparative Medicine, Yale University School of Medicine, New Haven, Connecticut, U.S.A. SfN 2011 – 351.22 Introduction Development of neurofibrillary tangles in Alzheimer's disease correlates with dementia and neuronal loss. Transgenic Tg4510 mice model tauopathy of the disease, and these mice exhibit progressive, region-specific neuron loss and behavioral deficits (Ramsden, 2005). The memory deficits associated with AD are thought to result largely from synaptic failure leading to hippocampal network dysfunction (Selkoe, 2002). Neurophysiological network activities, including theta rhythm, have been a major focus of studies of hippocampal function over decades (Buzsáki, 2002). Theta oscillation, an approximately sinusoidal (3 to 12 Hz) EEG activity that can be recorded in the limbic circuitry, has been linked to various cognitive processes and is considered to be a ‘natural tetanizer’ contributing to long-term potentiation (LTP) (Buzsáki, 2002; Vertes, 2005). One of the experimental models used to investigate hippocampal network activity in rodents involves electrical stimulation of key brain regions contributing to theta oscillation. High frequency stimulation of the brainstem nucleus pontisoralis (nPO) elicits robust hippocampal theta oscillation in anesthetized rats, where both the power and frequency have been shown to be stimulation current-dependent. Notably, it has been shown that a variety of drugs that disrupt cognitive functions, regardless of their synaptic mechanisms, diminish or eliminate elicited hippocampal theta oscillation (for review see (McNaughton et al., 2007)). Amyloid-β overproducing APP/PS1 mice have also shown a progressive decline in elicited hippocampal oscillations (Hajos et al. 2011) that correlates with deposition of amyloid plaques. In the present study, neuronal network activity in the limbic circuitry in 7 months old Tg4510 was investigated and compared to age matched wild-type mice. Multi-site field potentials were recorded using 16 channel NeuroNexus silicon probes, inserted across the hippocampus (CA1) and dentate gyrus (DG) in urethane anaesthetized mice. Population spikes in the DG were identified in response to perforant path stimulation. It was an essential step to identify locations of the recording sites across the hippocampal formation due to morphological changes in the hippocamus (cell degeneration and 50% loss of CA1 thickness) in Tg4510 mice, and to be able to compare electrophysiological signals between transgenic and WT mice. 7 month male tg4510 mice show decreased hippocampal volume, CA 1 pyramidal cell layer area and dentate gyrus granule cell body layer area tg4510 mice show deficits in perforant path evoked dentate gyrus population spike amplitude WT a. tg4510 b. *** 500µm 500µm ** *** c. d. *** Methods Summary of protocols used in the present study using urethane anaesthetized mice. A: Brainstem stimulation-evoked hippocampal LFP theta oscillation: the nPO was electrically stimulated and LFP in the hippocampal CA1 region was recorded. Following FFT of the LFP frequency of the highest power component (Peak fr.), and total power in the Θ frequency band (3 – 12 Hz) were recorded for each stimulation period (scale bars in the trace denote 1 sec time and 0.2 mV amplitude). B: Recording of population spikes and LTP: the perforant path (PP) was electrically stimulated using test stimuli and dentate gyrus (DG) response was recorded. Following baseline registration a theta burst protocol was used to induce LTP. Responses to test stimuli were recorded following LTP induction and increase of response amplitude compared to baseline was measured (scale bars in the trace denote 1 msec time and 0.5 mV amplitude; black trace denote recording closest to the DG granule cell layer). (Atlas images are modified from (Paxinos and Franklin, 2001)) Transgenic Animals: rTg4510P301L mice were produced by mating tetO-tau(P301L) Tg on a FVB/NTac background with CamKII-tTATg mice on a 129S6/SvEvTac background (Ramsden et al.) AT8 stained sections from 7 month male tg4510 mice and wild-type controls (*=P<0.05, **=P<0.01, ***=P<0.001 ; unpaired t-test) nPO evoked hippocampal oscillation is disrupted in 7 month male tg4510 mice Tg4510 mice show a reduction in maximal dentate gyrus population spike amplitude in response to perforant path stimulation. Stimulation-response curves from individual mice (a). While the maximal response produced is lower in tg4510 mice the current required to produce a population spike of 50% the amplitude of the maximal response is not significantly different (b). Amplitude and duration of LTP following theta burst stimulation is unchanged in tg4510 mice (c). Panel d shows mean average waveforms (n=5 / group) for evoked responses both pre-high frequency stimulation (30% of stimulation response curve maximum) and post theta burst stimulation. Tg4510 mice not only show a reduction in amplitude but also a change in the basic shape of the waveform, with very negligible, if any, excitatory post-synaptic potential (EPSP) observed. High frequency stimulation was able to potentiate the amplitude of population spike in both groups of animals (*=P<0.05, **=P<0.01, ***=P<0.001 ; unpaired t-test) Representative trace showing responses to 0.08 mA stimulation in WT and tg4510 mice. • Summary • 7 month male tg4510 mice show a significant reduction in overall hippocampal volume, including a drastic loss of cells in the CA1 pyramidal cell layer and the dentate gyrus cell body layer. • nPO stimulated hippocampal theta oscillations in tg4510 mice show hugely reduced total and theta power, although the frequency of response and the percentage of the total power in the theta band is unchanged. • Perforant path-evoked dentate gyrus population spike amplitude is drastically lower in tg4510 mice although both the magnitude and duration of LTP induced by theta burst stimulation is comparable to control animals. • Our electrophysiological findings are in line with histological observation of reduced cell number in the hippocampal formation and we further hypothesize sustained functionality of the surviving cells. nPO stimulated hippocampal theta oscillations in 7 month male tg4510 mice show no difference in peak frequency evoked or % power in theta range (3-12Hz) in response to varying stimulation intensities when compared to control mice. However, both total power and power in the theta band is significantly lower in the tg4510 mice when compared to their WT controls. (*=P<0.05, **=P<0.01, ***=P<0.001 ; two-way ANOVA) References Buzsáki G (2002) Theta oscillations in the hippocampus. Neuron 33:325-340. Hajos et al. 2011 Age-dependent disruption in hippocampal theta oscillation in Amyloid beta overproducing mice. Poster SfN 2011 241.21 McNaughton N, Kocsis B, Hajós M (2007) Elicited hippocampal theta rhythm: a screen for anxiolytic and procognitive drugs through changes in hippocampal function? Behav Pharmacol18:329-346. Ramsden et al. (2005) Age-Dependent Neurofibrillary Tangle Formation, Neuron Loss, and Memory Impairment in a Mouse Model of HumanTauopathy (P301L). J Neurosci 25: 10637-10647 Selkoe DJ (2002) Alzheimer's disease is a synaptic failure. Science 298:789-791. Vertes RP (2005) Hippocampal theta rhythm: a tag for short-term memory. Hippocampus 15:923-935. * *