Download
1 / 1
10 likes | 108 Views
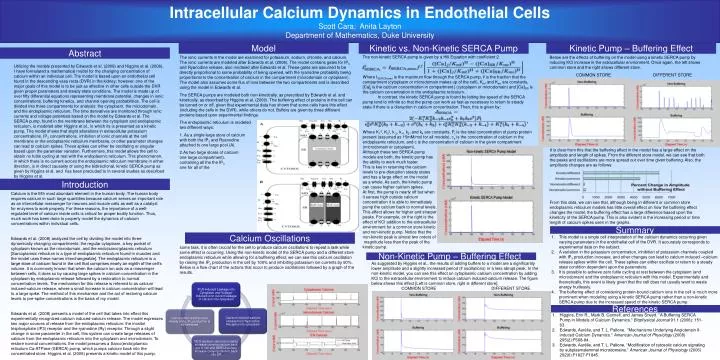
Intracellular Calcium Dynamics in Endothelial Cells Scott Cara; Anita Layton Department of Mathematics, Duke University. Model. Kinetic vs. Non-Kinetic SERCA Pump. Kinetic Pump – Buffering Effect. Abstract.

E N D
Intracellular Calcium Dynamics in Endothelial Cells Scott Cara; Anita Layton Department of Mathematics, Duke University Model Kinetic vs. Non-Kinetic SERCA Pump Kinetic Pump – Buffering Effect Abstract • The ionic currents in the model are examined for potassium, sodium, chloride, and calcium. The ionic currents are modeled after Edwards et al. (2008). The model contains gates for IP3 and Ryanodine release, also modeled after Edwards et al. These gates are assumed to be directly proportional to some probability of being opened, with the ryanodine probability being proportional to the concentration of calcium in the compartment (microdomain or cytoplasm). The model also assumes some flux of ions between the two compartments and is described using the model in Edwards et al. • The SERCA pumps are modeled both non-kinetically, as prescribed by Edwards et al. and kinetically, as described by Higgins et al. (2006). The buffering effect of proteins in the cell can be turned on or off, given that experimental data has shown that some cells have this effect (including the cells in the DVR), while others do not. Buffers are given by three different proteins based upon experimental findings. • The endoplasmic reticulum is modeled in two different ways: • As a single large store of calcium with both the IP3 and Ryanodine gates attached to one large pool (A). • As two large stores of calcium (in one large compartment), one containing all the the IP3 movement and one for all of the ryanodine receptor (B). • While acting as a cellular messenger to perform some task, it is often crucial for the cell to produce calcium oscillations to repeat a task while some effect is occurring. Using the non-kinetic model of the SERCA pump and a different store endoplasmic reticulum while allowing for a buffering effect, we can see this calcium oscillation by raising the IP3 production in the cell by 100% and inhibiting potassium ion currents by 90%. Below is a flow chart of the actions that occur to produce oscillations followed by a graph of the results. The non-kinetic SERCA pump is given by a Hill Equation with coefficient 2. Where ISERCA,max is the maximum flow through the SERCA pump, fj is the fraction that the compartment (cytoplasm or microdomain makes up of the cell), Kmf and Kmr are constants, [Ca]j is the calcium concentration in compartment j (cytoplasm or microdomain) and [Ca]SR is the calcium concentration in the endoplasmic reticulum. In contrast, the kinetic SERCA pump is found by letting the speed of the SERCA pump tend to infinite so that the pump can work as fast as necessary to return to steady state if there is a disruption in calcium concentration. Then, this is given by: Where K12, K32, k-2, k-4, k2, and k4 are constants, Pt is the total concentration of pump protein present (assumed as 15mM/mol for all models), ce is the concentration of calcium in the endoplasmic reticulum, and c is the concentration of calcium in the given compartment (microdomain or cytoplasm). Although these two SERCA pump models are both, the kinetic pump has the ability to work much faster. This is key in returning the calcium levels to pre-disruption steady states and has a large effect on the model as a whole. As such, the kinetic pump can cause higher calcium spikes. At first, the pump is nearly off but when it senses high outside calcium concentration it is able to immediately pump the calcium back to normal levels. This effect allows for higher and sharper peaks. For example, on the right is the effect of KCl addition to the extracellular environment for a common store kinetic and non-kinetic pump. Notice that the non-kinetic peak is almost two orders of magnitude less than the peak of the kinetic pump. Below are the effects of buffering on the model using a kinetic SERCA pump by inducing KCl increase in the extracellular environment. Once again, the left shows common store and the right shows different store. COMMON STORE DIFFERENT STORE It is clear from this that the buffering effect in the model has a large effect on the amplitude and length of spikes. From the different store model, we can see that both the peaks and oscillations are more spread out over time given buffering. Also, the amplitude changes are as follows: From this data, we can see that, although being in different or common store endoplasmic reticulum models has little overall effect on how the buffering effect changes the model, the buffering effect has a large difference based upon the kineticity of the SERCA pump. This is also evident in the increasing period or time length of calcium spikes seen in the graphs. Utilizing the models presented by Edwards et al. (2008) and Higgins et al. (2006), I have formulated a mathematical model for the changing concentration of calcium within an individual cell. The model is based upon an endothelial cell found in the descending vasa recta (DVR) in the kidney; however, one of the major goals of this model is to be just as effective in other cells outside the DVR given proper parameters and steady state conditions. The model is made up of over fifty differential equations representing membrane potential, changes in ionic concentrations, buffering kinetics, and channel opening probabilities. The cell is divided into three compartments for analysis: the cytoplasm, the microdomain, and the endoplasmic reticulum. The time derivatives are monitored through ionic currents and voltage potentials based on the model by Edwards et al. The SERCA pump, found in the membrane between the cytoplasm and endoplasmic reticulum, is modeled after Higgins et al., in which its is presented as a kinetic pump. The model shows that slight alterations in extracellular potassium concentrations, IP3 concentrations, inhibition of ionic channels at the cell membrane or the endoplasmic reticulum membrane, or other parameter changes can lead to calcium spikes. These spikes can either be oscillating or singular based upon the parameter variation. Furthermore, this model allows the cell to obtain no futile cycling at rest with the endoplasmic reticulum. This phenomenon, in which there is no current across the endoplasmic reticulum membrane in either direction, is in direct causality of using the bidirectional, kinetic SERCA pump as given by Higgins et al. and has been precluded to in several studies as described by Higgins et al. Introduction Calcium is the fifth most abundant element in the human body. The human body requires calcium in such large quantities because calcium serves an important role as an intercellular messenger for neurons and muscle cells as well as a catalyst for enzymes to work properly. For these reasons, the importance of a well-regulated level of calcium inside cells is critical for proper bodily function. Thus, much work has been done to properly model the dynamics of calcium concentrations within individual cells. Edwards et al. (2008) analyzed the cell by dividing the model into three dynamically changing compartments: the regular cytoplasm, a key pocket of cytoplasm known as the microdomain, and the endo(sarco)plasmic reticulum [Sarcoplamsic reticulum is a type of endoplasmic reticulum found in muscles and the model uses these names interchangeable]. The endoplasmic reticulum is a large store of calcium found in the cell that comprises nearly an eighth of the cell’s volume. It is commonly known that when the calcium ion acts as a messenger between cells, it does so by causing large spikes in calcium concentration in the cytoplasm by endoplasmic release followed by a restoration to normal concentration levels. The mechanism for this release is referred to as calcium induced-calcium release, where a small increase in calcium concentration will lead to a large spike. The method of this mechanism and the act of restoring calcium levels to pre-spike concentrations is the basis of my model. Edwards et al. (2008) presents a model of the cell that takes into effect this experimentally recognized calcium induced-calcium release. The model expresses two major sources of release from the endoplasmic reticulum: the inositol trisphosphate (IP3) receptor and the ryanodine (Ry) receptor. Through a slight change in some parameter in the cell, this system can create large releases of calcium from the endoplasmic reticulum into the cytoplasm and microdomain. To restore normal concentrations, the model presumes a Sarco(endo)plamsic reticulum Ca-ATPase (SERCA) pump, which pumps calcium back into the concentrated store. Higgins et al. (2006) presents a kinetic model of this pump. Summary Calcium Oscillations • This model is a single cell interpretation of the calcium dynamics occurring given varying parameters in the endothelial cell of the DVR. It accurately corresponds to experimental data on the subject. • A variation in the potassium concentration, inhibition of potassium channels coupled with IP3 production increase, and other changes can lead to calcium induced –calcium release spikes within the cell. These spikes can either oscillate or return to a steady state condition dependent upon the parameters. • It is possible to achieve zero futile cycling at rest between the cytoplasm (and microdomain) and the endoplasmic reticulum with this model. Experimentally and theoretically, this event is likely given that the cell does not usually want to waste energy fruitlessly. • The buffering effect of considering protein-bound calcium ions in the cell is much more prominent when modeling using a kinetic SERCA pump rather than a non-kinetic SERCA pump due to the increased speed of the kinetic SERCA pump. Non-Kinetic Pump – Buffering Effect As suggested by Higgins et al., the results of adding buffers to a model are a significantly lower amplitude and a slightly increased period (if oscillations) or a less abrupt peak. In the non-kinetic model, you can see this effect on cytoplasmic calcium concentration by adding KCl to the extracellular environment to induce calcium induced-calcium release. The figure below shows this effect [Left is common store, right is different store]. COMMON STORE DIFFERENT STORE References • Higgins, Erin R., Mark B. Cannell, and James Sneyd. "A Buffering SERCA Pump in Models of Calcium Dynamics." Biophysical Journal 91.1 (2006): 151-63. • Edwards, Aurélie, and T. L. Pallone. "Mechanisms Underlying Angiotensin II-induced Calcium Dynamics." American Journal of Physiology (2008) 295(2):F568-84. • Edwards, Aurélie, and T. L. Pallone. “Modification of cytosolic calcium signaling by subplasmalemmal microdomains.” American Journal of Physiology (2006) 292(6):F1827-F1845.