Download
1 / 1
10 likes | 122 Views
Quantification of magnetization transfer and relaxation rates by MT-prepared multi-echo EPI Gunther Helms 1,2 , Gisela E. Hagberg 3 1 MR-Research in Neurology and Psychiatry, Faculty of Medicine, University of Göttingen, D-37075 Germany

E N D
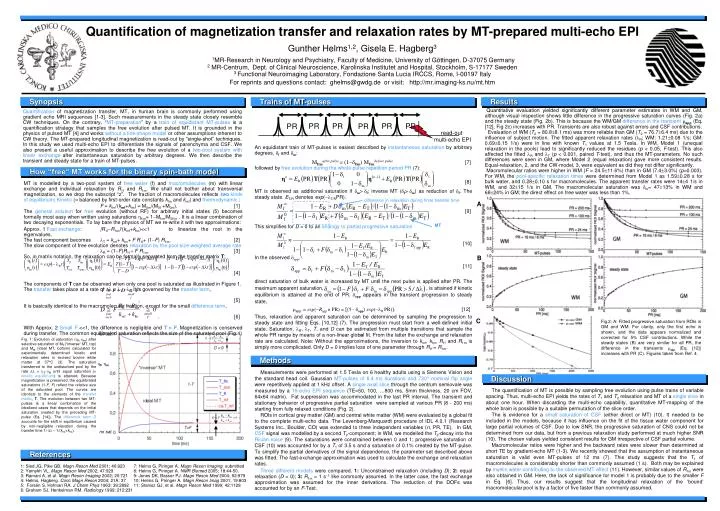
Quantification of magnetization transfer and relaxation rates by MT-prepared multi-echo EPI Gunther Helms1,2, Gisela E. Hagberg3 1MR-Research in Neurology and Psychiatry, Faculty of Medicine, University of Göttingen, D-37075 Germany 2 MR-Centrum, Dept. of Clinical Neuroscience, Karolinska Institutet and Hospital, Stockholm, S-17177 Sweden 3 Functional Neuroimaging Laboratory, Fondazione Santa Lucia IRCCS, Rome, I-00197 Italy For reprints and questions contact: ghelms@gwdg.de or visit: http://mr.imaging-ks.nu/mt.htm Synopsis Trains of MT-pulses Results Quantitative evaluation yielded significantly different parameter estimates in WM and GM, although visual inspection shows little difference in the progressive saturation curves (Fig. 2a) and the steady state (Fig. 2b). This is because the WM/GM difference in the transientmapp (Eq. [12], Fig 2c) increases with PR. Transients are also robust against errors and CSF contributions. Evaluation of WM (T2 = 80.88.1 ms) was more reliable than GM (T2 = 76.76.4 ms) due to the influence of subject motion. The fitted apparent relaxation rates (lR; WM: 1.210.08 1/s; GM: 0.690.15 1/s) were in line with known T1 values at 1.5 Tesla. In WM, Model 1 (unequal relaxation in the pools) lead to significantly reduced the residues (p < 0.05, F-test). This also affected the fitted lR and lT (p < 0.001, paired T-test), and thus the MT-parameters. No such differences were seen in GM, where Model 2 (equal relaxation) gave more consistent results. Equal-relaxation, 2, and the CW-model, 3, were equivalent as did they not differ significantly. Macromolecular ratios were higher in WM (F = 24.511.6%) than in GM (7.43.0%) (p=0.003). For WM, the pool-specific relaxation times were determined from Model 1 as 1.590.28 s for bulk water, and 0.280.10 s for macromolecules. The apparent transfer rates were 144 1/s in WM, and 3215 1/s in GM. The macromolecular saturation was dm= 4713% in WM and 6824% in GM; the direct effect on free water was less than 1%. Quantification of magnetization transfer, MT, in human brain is commonly performed using gradient echo MRI sequences [1-3]. Such measurements in the steady state closely resemble CW techniques. On the contrary, “MT-preparation” by a train of equidistant MT-pulses is a quantification strategy that samples the free evolution after pulsed MT. It is grounded in the physics of pulsed MT [4] and works without a line-shape model or other assumptions inherent to CW theory. The MT-prepared longitudinal magnetization is read-out by “single-shot” techniques. In this study we used multi-echo EPI to differentiate the signals of parenchyma and CSF. We also present a useful approximation to describe the free evolution of a two-pool system with linear exchangeafter instantaneous saturation by arbitrary degrees. We then describe the transient and steady state for a train of MT-pulses. An equidistant train of MT-pulses is easiest described by instantaneous saturationby arbitrary degrees,df and dm: Mf/mafter pulse = (1-df/m) Mf/mbefore pulse [7] followed by free evolution during the whole pulse repetition period PR (7): [8] MT is observed as additional saturation if dm>df; inverse MT (df>dm) as reduction of df. The steady state. ET/R denotes exp(-lT/RPR). [9] This simplifies for D = 0 to an analogy to partial progressive saturation [10] In the observed dapp, [11] direct saturation of bulk water is increased by MT until the next pulse is applied after PR. The maximum apparent saturation, , is attained if kinetic equilibrium is attained at the end of PR. dapp appears in the transient progression to steady state, mapp = exp(-Rappn PR) = [(1-dapp) exp(-lR PR)]. [12] Thus, relaxation and apparent saturation can be determined by sampling the progression to steady state and fitting Eqs. [10,12] (7). The progression must start from a well-defined initial state. Saturation, lR, lT, T, and D can be estimated from multiple transitions that sample the whole PR range by means of a non-linear global fit. From the latter the exchange and relaxation rate are calculated. Note: Without the approximations, the inversion to kmf, kfm, R1f and R1m is simply more complicated. Only D = 0 implies loss of one parameter through R1f = R1m. PR PR PR PR PR PR How “free“ MT works for the binary spin-bath model MT is modelled by a two-pool system of free water (f) and macromolecules (m) with linear exchange and individual relaxation by R1f and R1m. We shall not bother about transversal magnetization, so we drop the subscript “z”. The fraction of macromolecules reflects two kinds of equilibrium: Kinetic (= balanced by first-order rate constants kfm and kmf) and thermodynamic: F = kfm/(kmf+kfm) = M0m/(M0f +M0m). [1] The general solution for free evolution (without RF) for arbitrary initial states (5) becomes formally most easywhen written using saturations hf/m= 1-Mf/m/M0f/m . It is a linear combination of two decaying exponentials. To lay bare the physics of MT we re-write it with two approximations: Approx. 1 Fast exchange: |R1f-R1m|/(kmf+kfm)<<1 to linearize the root in the eigenvalues. The fast component becomes lT = kmf+ kfm+ FR1f+ (1-F) R1m. [2] The slow component of free evolution denotes relaxation by the pool-size-weighted average rate lR = (1-F)R1f + F R1m. [3] So, in matrix notation, the relaxation can be formally separated from the transfer matrix T: [4] The components of T can be observed when only one pool is saturated as illustrated in Figure 1. The transfer takes place at a rate of Dl = lT-lR It is governed by the transfer term, , [5] It is basically identical to the macromolecular fraction, except for the small difference term, . [6] With Approx. 2 Small F <<1, the difference is negligible and T = F. Magnetization is conserved during transfer. The common equilibrated saturation reflects the size of the saturated pool (Fig.1) read-out multi-echo EPI difference in relaxation during finite transfer time A MT B C Fig.2: A: Fitted progressive saturation from ROIs in GM and WM. For clarity, only the first echo is shown, and the data appears normalized and corrected for 5% CSF contributions. While the steady states (B) are very similar for all PR, the difference in the transients mapp (Eq. [12]) increases with PR (C). Figures taken from Ref. 4. full sat. n Fig. 1: Evolution of saturation (hf, hm) after selective saturation of Mf (“inverse“ MT, top) and Mm (ideal MT, bottom) calculated for experimentally determined kinetic and relaxation rates in excised bovine white matter at 37°C (3). The saturation transferred to the undisturbed pool by the rate Dl = lT-lR until equal saturation(= kinetic equilibrium) is attained. Because magnetization is preserved, the equilibrated saturations (1-F, F) reflect the relative size of the saturated pool. The curves are identical to the elements of the transfer matrix, T. The evolution between two MT-pulses is a linear combination of the idealized cases that depends on the initial saturation created by the preceding MT-pulse (Eq. [14]). The difference term D accounts for the shift in equilibrium caused by non-negligible relaxation during the transfer time 1/ Dl≈ 1/(kmf+kfm). Methods hf, hm Measurements were performed at 1.5 Tesla on 6 healthy adults using a Siemens Vision and the standard head coil. Gaussian MT-pulses of 6.4 ms durations and 720° nominal flip angle were repetitively applied at 1 kHz offset. A single axial slice through the centrum semiovale was measured by a 16-echo EPI sequence (TE=50, 100, …800 ms, 5mm thickness, 20 cm FOV, 64x64 matrix). Fat suppression was accommodated in the last PR interval.The transient and stationary behavior of progressive partial saturation were sampled at various PR (8 - 200 ms) starting from fully relaxed conditions (Fig. 2). ROIs in cortical grey matter (GM) and central white matter (WM) were evaluated by a global fit to the complete multi-echo data. The Levenberg-Marquardt procedure of IDL 4.0.1 (Research Systems Inc., Boulder, CO) was extended to three independent variables (n, PR, TE). In GM, CSF signal was modelled by a second T2-component; in WM, we modelled the T2-decay into the Rician noise (9). The saturations were constrained between 0 and 1; progressive saturation of CSF (10) was accounted for by a T1 of 3.5 s and a saturation of 0.1% created by the MT-pulse. To simplify the partial derivatives of the signal dependence, the parameter set described above was fitted. The fast-exchange approximation was used to calculate the exchange and relaxation rates. Three different models were compared. 1: Unconstrained relaxation (including D); 2: equal relaxation (D = 0); 3:R1m = 1 s-1 like commonly assumed. In the latter case, the fast exchange approximation was assumed for the inner derivations. The reduction of the DOFs was accounted for by an F-Test. Discussion The quantification of MT is possible by sampling free evolution using pulse trains of variable spacing. Thus, multi-echo EPI yields the rates of T1 and T2 relaxation and MT of a single slice in about one hour. When discarding the multi-echo capability, quantitative MT-mapping of the whole brain is possible by a suitable permutation of the slice order. The is evidence for a small saturation of CSF (either direct or MT) (10). It needed to be included in the models, because it has influence on the fit of the tissue water component for large partial volumes of CSF. Due to low SNR, the progressive saturation of CNS could not be determined from our data, but from a previous relaxation study performed at much higher SNR (10). The chosen values yielded consistent results for GM irrespective of CSF partial volume. Macromolecular ratios were higher and the backward rates were slower than determined at short TE by gradient-echo MT (1-3). We recently showed that the assumption of instantaneous saturation is valid even MT-pulses of 12 ms (7). This study suggests that the T1 of macromolecules is considerably shorter than commonly assumed (1 s). Both may be explained by myelin water contributing to the observed MT effect (11). However, similar values of R1m were also obtained in GM. Here, the lack of significance for model 1 is probably due to the smaller F in Eq. [6]. Thus, our results suggest that the longitudinal relaxation of the ‘bound’ macromolecular pool is by a factor of five faster than commonly assumed. no sat. References 1: Sled JG, Pike GB. Magn Reson Med 2001; 46:923 2: Yarnykh VL. Magn Reson Med 2002; 47:929 3: Ramani A, et al. Magn Reson Imaging 2002; 20:721 4: Helms, Hagberg. Conc Magn Reson 2004; 21A: 37 5: Forsén S, Hofman RA. J Chem Phys 1963; 39:2892 6: Graham SJ, Henkelman RM. Radiology 1999; 212:231 7: Helms G, Piringer A. Magn Reson Imaging; submitted 8: Helms G, Piringer A. NMR Biomed 2005; 18:44-50. 9: Jones DK, Basser PJ. Magn Reson Med 2004; 52:979 10: Helms G, Piringer A. Magn Reson Imag 2001; 19:803 11: Stanisz GJ, et al. Magn Reson Med 1999; 42:1128