Download

1 / 27
280 likes | 383 Views
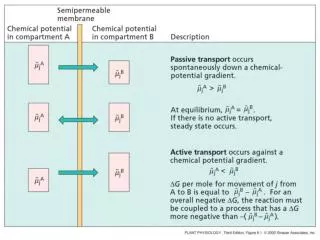
The (resting) voltage difference from inside a (typical) neuron to “outside” the cell membrane is about -70mv Resting potential due to sodium ions being pumped out and potassium ions being pumped in. At rest potassium conductance greater than sodium’s

E N D
The (resting) voltage difference from inside a (typical) neuron to “outside” the cell membrane is about -70mv • Resting potential due to sodium ions being pumped out and potassium ions being pumped in. • At rest potassium conductance greater than sodium’s • Balance diffusion current flux with current due to drift in E field across membrane to calculate Nernst potential • Compare to thermocouples, where two metals, dissimilar in mobility, are welded together and their interface is able to generate a microvolt-size potential difference. Nernst Potential
Transport • A membrane protein, Na+/K+ ATPase: moves 3 Na+ out and 2 K+ in… 3 vs 2? • Until proper concentration gradient achieved: (membrane) voltage-controlled pump • Energy comes from the hydrolysis of ATP
From Gary G. Matthews, Cellular Physiology of Nerve and Muscle (1991) A- represents negatively charged protein
Membrane channels www.neurosearch.com/
Reading: George B. Benedek & Felix M. H. Villars, Physics With Illustrative Examples from Medicine and Biology, Vol 3: Electricity and Magnetism, Addison-Wesley (1979). Howard C. Berg, Random Walks in Biology, Princeton Univ. Press (1983). A statistical physics look at the diffusion-drift development that leads to the Nernst potential. Berg is well-known for his "Life at low Reynolds number" essay: see p. 75 of the book. Bertil Hille, Ion Channels of Excitable Membranes 3rd Ed, Sinauer & Associates, 814 pp. (2001)
Flux as current: • Units of D: cm^2/sec. • Fick's 1st Law tells us about a diffusive flux of particles, charged or uncharged. • For example, glucose is an uncharged particle in solution, and is subject to Fick's Law just as well as charged K+ and Cl-, but glucose flux is NOT a current! • A flux of sodium cations (+) is a positive current in the same direction as the flux, while a flux of chloride anions (-) is a current in the opposite direction. • The diffusion of charged particles (in the case we‘ll considering, of K+) will set up an E field which opposes the diffusion flux. • When the two fluxes are equal and opposite the voltage difference across the membrane will be at equilibrium • Just how does an ion diffuse across a lipid bilayer?
Drift of charged particles in an E field: • In a material, charged particles will "drift" with a velocity proportional to their mobility μ, their charge, and the strength of the E field: • If the charged particles were a plasma in a vacuum they would move under the influence of F=ma, accelerating, but here in a material the particles reach a "terminal drift velocity". • ~drift in gravitational field… Demo with corn syrup
Flux as current: from we find the drift flux is • Where C(x) is the concentration of the ion in question • units of mobility μ are cm^2/(volt-sec-coulomb) • corn syrup demo: steel 7.8, glass 2.4, Al 2.5 spec den
We can now algebraically add the diffusion and drift flux in a steady-state version of KCL What is the relationship between mobility μand diffusion factor D? Einstein found: (in metals), where gas constant R and Boltzmann constant k are related by R = kN, where N is the number of molecules in question. (ref: Van Vlack, Elements of Materials Science 2nd Ed., 1964. pp 105, 98). As a result (considering other material factors too), when temperature rises, diffusion coefficient D always increases while mobility μ increases for non-metals and decreases for metals. Further information in: F. Reif, Statistical Physics (1967), page 337 shows that, in general, Viscosity?
Now we need to remind you that Since this integral is "conservative" we can go along any path from “gnd” to point P (in our case, from outside to inside the cell, across the membrane). Integrate the flux balance equation to end up computing voltage.
Remember log(X) - log(Y) = log(X/Y), and know that at room temperature, kT/q = 25 mV we have the Nernst equation for a singly-charged positive ionic species at room temp. Consider a ratio of internal to external potassium of 10:1, we find that VK = -58 mV, which turns out to be what is measured.
Given the concentration gradient of sodium Na+, what will be the sign of the "sodium equilibrium potential" (Nernst potential considering sodium alone)? Because sodium concentration is higher outside the cell than in, its Nernst potential will be positive, and will follow the same Nernst equation logarithmic law as potassium.
What about calcium as Ca++? • Ca++ is a doubly charged ion. Therefore substitute 2q in the kT/”q” term of the Nernst equation. • The Nernst voltage is reduced by a factor of 2! • Think of it this way: in the same E field a Ca++ ion will experience twice the force as a K+ ion. Therefore half the field strength would be needed to exert the same force on Ca++.
What about chloride ions? • There are two thoughts to have about chloride: • (1) Since chloride ion has the opposite sign of K, then all other things being equal, the sign of the answer for the Nernst potential should be opposite to that of potassium. • (2) Since chloride actually has a higher concentration outside than inside the cell, then the sign of the answer should be the same as potassium.
chloride drift • Consider the drift flux of Cl- due to the electric field of charge separation. In our K+ equation the term q was +e, where e is the magnitude of the charge on an electron. Now q becomes -e for the chloride ion. But the electric field changes direction too, because negative charges instead of positive charges have moved into position to block the further diffusion of chloride ions.
Two ionic species, same concentrations, but unequal mobility factors μ: • Say the concentration of K+ and Cl- is the same C(x) everywhere and that the mobility of K+ > mobility Cl-. What is an expression for the transmembrane voltage? Potassium flux: Chloride flux: Note sign of electric field drift is same in both expressionsand C(x) is the same for both…
now integrate from OUT to IN, as before, and obtain Where μ no longer cancels out;if μCl = 0 then the previous form appears
generalizing further, and taking account ofdifferent ionic concentrations: where N is the total number of ionic species, and Vj is the Nernst potential of the jth species. Example where 2 concentrations are different. Assume the concentration ratios of Na and K are (1/10) and (20/1) respectively and the mobility of K is 3x more than Na. What is the equilibrium potential? VK = -25*ln(20) = -75mv; VNa = +25*ln(10) = 58mvVINSIDE = -75*(3/4) + 58*(1/4) = -39 mV (weighted sum)
Nature of the channel • On a macroscopic level the current crossing a membrane through channels can seem to vary continuously… • On a micro level individual channels go from open to closed and back with v. little time in transition…
Membrane conductance as a function of temperature • We saw differences between semiconductor and metal conductance as a function of temperature… • So far we know about membrane voltage change as a function of temperature…what about current (conductance)? • From Hille, Ionic Channels of Excitable Membranes: “Warming an axon by 10deg C speeds up the rates of gating 2-4 fold (Q10). As we know now, gating involves conformational changes of channel proteins, and the rates of these conformation changes are temperature-sensitive.” • “In the range between 19° C and 37° C, the membrane conductance increased with rising temperature and decreased with cooling (Mean Q10 about 2·5).” Alison Brading, Edith Bülbring, and T. Tomita 1969 • And we have seen specific temperature-sensitive membrane proteins…in epidermis and hypothalamus…
Temperature Dependence of Ion Permeation at the Endplate Channel HELENE M. HOFFMANN and VINCENT E. DIONNE (1983) From the Department of Medicine, Division of Pharmacology, UCSD ABSTRACT The dependence of acetylcholine receptor mean single-channel conductance on temperature was studied at garter snake twitch-muscle endplates using fluctuation analysis. In normal saline under conditions where most of the endplate current was carried by Na +, the channel conductance increased continuously from near 0°C to ~23°C with a Q10 of 1.97 (4- 0.14) (mean + SD). When 50% of the bath Na + was replaced by either Li +, Rb +, or Cs +, the Q10 did not change significantly; however, at any temperature the channel conductance was greatest in Cs-saline and decreased with the ion sequence Cs > Rb > Na > Li. The results were fit by an Eyring-type model consisting of one free-energy well on the extracellular side of a single energy barrier. Ion selectivity appeared to result from ion-specific differences in the well and not in the barrier of this model. With a constant barrier enthalpy for different ions, well free-energy depth was greatest for Cs + and graded identical to the permeability sequence. The correlation between increased well depth (i.e., ion binding) and increased channel conductance can be accounted for by the Boltzmann distribution of thermal energy.
Action Potential • When a nerve or muscle cell is stimulated by synaptic transmission, the mobility (or channel conductance, or permeability) for sodium transiently increases to a value greater than potassium’s and the cell’s internal voltage "spikes" above zero for about a millisecond. www.bio.psu.edu/Courses/Fall2002/Biol142/neurons/neurons.html
AP propagation speeds • 5-120 m/sec range • Compare to 10^8 m/s speed of light (electronics) • Speed proportional to (sqrt) axon diameter • “The speed of propagation for mammalian motor neurons is 10 - 120 m/s, while for nonmyelinated sensory neurons it's about 5 - 25 m/s” • www.rwc.uc.edu/koehler/biophys/4d.html • http://www.youtube.com/watch?v=Jo8rOHEHy4wTarantula vs prey
Patch clamping channels for ions • Most ions move relatively freely inside and outside the cells; it's at the membrane barrier that mobility becomes important • Proteins form channels for specific ions • Permeability (a more common term for ionic mobility in the membrane) of a channel can be modulated by synaptic activity or transmembrane voltage. • It is possible to record the current passing through individual channels with a patch clamp electrode isolating a small section of membrane
1991 Nobel Prize in Physiology or Medicine to Bert Sakmann and Erwin Neher from Germany for their work on patch clamping