Download
1 / 1
10 likes | 128 Views
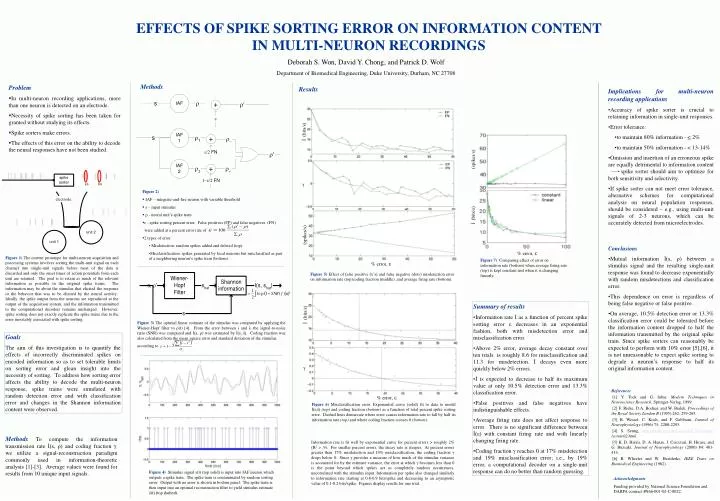
Implications for multi-neuron recording applications Accuracy of spike sorter is crucial to retaining information in single-unit responses. Error tolerance: to maintain 80% information - < 2% to maintain 50% information - < 13-14%

E N D
Implications for multi-neuron recording applications • Accuracy of spike sorter is crucial to retaining information in single-unit responses. • Error tolerance: • to maintain 80% information - < 2% • to maintain 50% information - < 13-14% • Omission and insertion of an erroneous spike are equally detrimental to information content spike sorter should aim to optimize for both sensitivity and selectivity. • If spike sorter can not meet error tolerance, alternative schemes for computational analysis on neural population responses, should be considered – e.g., using multi-unit signals of 2-3 neurons, which can be accurately detected from microelectrodes. • Conclusions • Mutual information İ(s, ρ)between a stimulus signal and the resulting single-unit response was found to decrease exponentially with random misdetections and classification error. • This dependence on error is regardless of being false negative or false positive. • On average, 10.5% detection error or 13.3% classification error could be tolerated before the information content dropped to half the information transmitted by the original spike train. Since spike sorters can reasonably be expected to perform with 10% error [5],[6], it is not unreasonable to expect spike sorting to degrade a neuron’s response to half its original information content. İ (bits/s) (spikes/s) γ • Figure 2) • IAF – integrate-and-fire neuron with variable threshold • s – input stimulus • r - neural unit’s spike train. • ε - spike sorting percent error. False positives (FP) and false negatives (FN) were added at a percent error rate of • 2 types of error • Misdetection: random spikes added and deleted (top). • Misclassification: spikes generated by local neurons but misclassified as part of a neighboring neuron’s spike train (bottom). % error, ε Figure 6) Misclassification error. Exponential curve (solid) fit to data to model İ(s,ŝ) (top) and coding fraction (bottom) as a function of total percent spike sorting error. Dashed lines demarcate where error causes information rate to fall by half its information rate (top) and where coding fraction crosses 0 (bottom). (spikes/s) % error, ε Figure 7) Comparing effect of error on information rate (bottom) when average firing rate (top) is kept constant and when it is changing linearly. % error, ε Figure 5)Effect of false positive (x’s) and false negative (dots) misdetection error on information rate (top)coding fraction (middle), and average firing rate (bottom). . Figure 3) The optimal linear estimate of the stimulus was computed by applying the Wiener-Hopf filter to ρ(t) [4]. From the error between s and ŝ, the signal-to-noise ratio (SNR) was computed and İ(s, ρ) was estimated by İ(s, ŝ). Coding fraction was also calculated from the mean square error and standard deviation of the stimulus, according to . İ (bits/s) Figure 4) Stimulus signal s(t) (top solid) is input into IAF neuron which outputs a spike train. The spike train is contaminated by random sorting error. Output with no error is shown in bottom panel. The spike train is then input into an optimal reconstruction filter to yield stimulus estimate ŝ(t) (top dashed). Deborah S. Won, David Y. Chong, and Patrick D. Wolf Department of Biomedical Engineering, Duke University, Durham, NC 27708 EFFECTS OF SPIKE SORTING ERROR ON INFORMATION CONTENT IN MULTI-NEURON RECORDINGS Methods • Problem • In multi-neuron recording applications, more than one neuron is detected on an electrode. • Necessity of spike sorting has been taken for granted without studying its effects. • Spike sorters make errors. • The effects of this error on the ability to decode the neural responses have not been studied. Results electrode Figure 1) The current prototype for multi-neuron acquisition and processing systems involves sorting the multi-unit signal on each channel into single-unit signals before most of the data is discarded and only the onset times of action potentials from each unit are retained. The goal is to retain as much of the relevant information as possible in the original spike trains. The information may be about the stimulus that elicited the response or the behavior that was to be elicited by the neural activity.. Ideally, the spike output from the neurons are reproduced at the output of the acquisition system, and the information transmitted to the computational decoders remains unchanged. However, spike sorting does not exactly replicate the spike trains due to the error inevitably associated with spike sorting. • Summary of results • Information rate İ as a function of percent spike sorting error εdecreases in an exponential fashion, both with misdetection error and misclassification error. • Above 2% error, average decay constant over ten trials is roughly 8.6 for misclassification and 11.3 for misdetection. İ decays even more quickly below 2% errors. • İ is expected to decrease to half its maximum value at only 10.5% detection error and 13.3% classification error. • False positives and false negatives have indistinguishable effects. • Average firing rate does not affect response to error. There is no significant difference between İ(ε) with constant firing rate and with linearly changing firing rate. • Coding fraction γ reaches 0 at 17% misdetection and 19% misclassification error; i.e., by 19% error, a computational decoder on a single-unit response can do no better than random guessing. İ (bits/s) Goals The aim of this investigation is to quantify the effects of incorrectly discriminated spikes on encoded information so as to set tolerable limits on sorting error and glean insight into the necessity of sorting. To address how sorting error affects the ability to decode the multi-neuron response, spike trains were simulated with random detection error and with classification error and changes in the Shannon information content were observed. γ References [1] Y. Tock and G. Inbar, Modern Techniques in Neuroscience Research, Springer-Verlag, 1999. [2] F. Rieke, D.A. Bodnar, and W. Bialek, Proceedings of the Royal Society London B (1995) 262: 259-265. [3] R. Wessel, C. Koch, and F. Gabbiani, Journal of Neurophysiology (1996) 75: 2280-2293. [4] S. Seung, http://hebb.mit.edu/courses/9.29/lectures/ lecture02.html. [5] K. D. Harris, D. A. Henze, J. Csicsvari, H. Hirase, and G. Buzsaki, Journal of Neurophysiology (2000) 84: 401-414. [6] B. Wheeler and W. Heetderks, IEEE Trans on Biomedical Engineering (1982). Methods To compute the information transmission rate İ(s, ρ) and coding fraction γ, we utilize a signal-reconstruction paradigm commonly used in information-theoretic analysis [1]-[3]. Average values were found for results from 10 unique input signals. Information rate is fit well by exponential curve for percent errors > roughly 2% (R2 > .9). For smaller percent errors, the decay rate is steeper. At percent errors greater than 17% misdetection and 19% misclassification, the coding fraction γ drops below 0. Since γ provides a measure of how much of the stimulus variance is accounted for by the estimate variance, the error at which γbecomes less than 0 is the point beyond which spikes act as completely random occurrences, uncorrelated with the stimulus input. Information per spike also changed similarly to information rate starting at 0.8-0.9 bits/spike and decreasing to an asymptotic value of 0.1-0.2 bits/spike. Figures display results for one trial. Acknowledgments Funding provided by National Science Foundation and DARPA contract #N66-001-02-C-8022.