Download

1 / 36
370 likes | 606 Views
Section Q Protein Synthesis. Q1 Aspects of Protein Q2 Mechanism of Protein Synthesis Q3 Initiation in Eukaryotes Q4 Translation Control and Post-translational Events. Q1 Aspects of Protein. Codon-anticodon interaction Wobble Ribosome binding site Polysomes Initiator tRNA.

E N D
Section Q Protein Synthesis Q1 Aspects of Protein Q2 Mechanism of Protein Synthesis Q3 Initiation in Eukaryotes Q4 Translation Control and Post-translational Events Yang Xu, College of Life Sciences
Q1 Aspects of Protein • Codon-anticodon interaction • Wobble • Ribosome binding site • Polysomes • Initiator tRNA Yang Xu, College of Life Sciences
Codon-anticodon Interaction In the cleft of the ribosome, an anti-parallel formation of three base pairs occurs between the codon on the mRNA and the anticodon on the tRNA. The anticondon at one end of the tRNA interacts with a complementary triplet codon of bases on the mRNA, when both are brought together in the cleft of the ribosome. The interaction is anti-parallel in nature. Some highly purified tRNA molecules were found to interact with more than one codon, and this ability correlated with the presence of modified nucleosides in the 5’-anticodon position. Yang Xu, College of Life Sciences
Wobble-I • The wobble hypothesis was suggested by Francis Crick to explain the redundancy of genetic code. • His specific predictions are shown in right table along with actual observation. Yang Xu, College of Life Sciences
Wobble-II To explain the redundancy of the genetic code. 18 aa are encoded by more than one triplet codons which usually differ at 5’-anticodin base. The 5'-anticodon base is able to undergo more movement than the other two bases and can thus form non-standard base pairs as long as the distances between the ribose units are close to normal. Yang Xu, College of Life Sciences
Wobble-III Base pairings at the wobble position I: inosine Yang Xu, College of Life Sciences
Ribosome mRNA Ribosome binding site- Shine-Dalgarno sequence • Only for prokaryotic translation • A purine-rich sequence usually containing all or part of the sequence 5'-AGGAGGU-3' • Locates 8-13 nt upstream of the initiation codon in prokaryotic mRNA • Function: To position the ribosome binding for initiation of protein synthesis Ribosome Yang Xu, College of Life Sciences
Ribosome binding site- Shine-Dalgarno sequence • John Shine is known to most undergraduate biology students for his role in defining the Shine-Dalgarno gene sequence, which is responsible for the initiation and termination of protein-synthesis. Education 1972 BSc (Hons) 1st Class, Australian National University 1975 PhD, Australian National University 2006 DSc (Honoris causa) The University of New South Yang Xu, College of Life Sciences
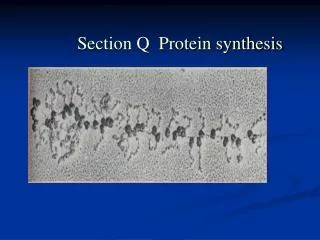
Polysomes • Each mRNA transcript is read by more than one ribosome. • A second, third, fourth, etc. ribosome starts to read the mRNA transcript before the first ribosome has completed the synthesis of one polypeptide chain. • Multiple ribosomes on a single mRNA transcript are called polyribosomes or polysomes. • Multiple ribosomes can not be positioned closer than 80 nt. Polysomes Yang Xu, College of Life Sciences
Polysomes • Electron micrographs of ribosomes actively engaged in protein synthesis revealed by "beads on a string" appearance. Yang Xu, College of Life Sciences
fMet-tRNAfMet Initiator tRNA The initiator tRNA fMet-tRNAfMet Yang Xu, College of Life Sciences
Q2 Mechanism of Protein Synthesis • Overview • Initiation • Elongation • Termination Yang Xu, College of Life Sciences
Overview Three stages: • Initiation: the assembly of a ribosome on an mRNA molecule; • Elongation: repeated cycles of amino acid addition; • Termination: the release of the new protein chain. Yang Xu, College of Life Sciences
Initiation factors 30S subunit 1 2 3 G G G 2 2 Shine-Dalgarno sequence 1 3 fMet mRNA AUG 3 Initiator tRNA fMet 1 30S initiation Complex 3’ 3’ G 5’ 5’ 2 1 3’ 50S subunit 5’ P-site A-site 70S initiation Complex Initiation in E coli Initiation Steps 1. IF1 and IF3 bind to a free 30S subunit. 2. IF2 complexed with GTP then binds to the small subunit. It will assist the charged initiation tRNA to bind. 3. The 30S subunit attaches to an mRNA molecule marking use of the ribosome-binding site (RBS) on the mRNA. 4. The initiator tRNA can then bind to the complex by base pairing of its anti-codon with the mRNA. 5. The 50S subunit can now bind, which displaces IF1 and IF2, and the GTP is hydrolyzed in this energy-consuming step. 6. 70S initiation complex formed at the end of this phase. Yang Xu, College of Life Sciences
P-site Empty A-site aa aa aa aa aa aa aa aa Binding of aa-tRNA to A-site 3’ Peptide bond formation 5’ 3’ 3’ 3’ aa aa aa aa From A- to P-site 5’ 5’ 5’ Elongation in E coli Elongation Steps 1. Aminoacyl-tRNA delivery. EF-Tu is required to delivery the aa-tRNA to A-site. Energy is from GTP. 2. Peptide bond formation. After aa-tRNA delivery, the A- and P-site are both occupied and the two amino acids that are joined closely. The peptidyl transferase activity of the 50S subunit can now form a peptide bond between these two amino acids. 3. Translocation. A complex of EF-G (translocase) and GTP binds to the ribosome and the discharged tRNA is elected from the P-site, the peptidyl-tRNA is moved from the A-site to P-site. Yang Xu, College of Life Sciences
EF-G:GDP Aa-tRNA Aa-tRNA-EF-Tu:GTP EF-Ts EF-Tu:GTP GTP EF-Tu:EF-Ts EF-G:GTP EF-Tu:GDP GDP EF-Ts Yang Xu, College of Life Sciences
Role of elongation factors in prokaryotes • EF-Tu • It is a G protein (GTP/GDP-binding protein) • Active when GTP is bound (Kd = 10-6 M) • Inactive with GDP is bound (Kd = 10-8 M), slow offrate • Its off rate is increased by EF-Ts • EF-Ts • It is a guanosine nucleotide exchange factor (GEF) for EF-Tu • EF-G • It is a G protein • There is no known GEF for EF-G Yang Xu, College of Life Sciences
N Termination Termination codon UGA • There are no tRNA species that normally recognize stop codon. • Instead, protein factors called release factors interact with these codons. RF1 recognizes UAA and UAG, and RF2 recognizes UAA and UGA. • RF3 helps either RF1 or RF2 to carry out the reaction. • The release factors mark peptidyl transferase transfer polypeptide to water rather than to the usual aminoacyl-tRNA, and thus the new protein is released. • To remove the uncharged tRNA from P-site and release the mRNA, EF-G is needed. Release factor 1 or 2 G aa aa Release factor 3 aa aa G Peptidyl transerase Released polypeptide chain G aa aa aa aa aa aa aa aa aa aa aa aa aa aa aa aa aa aa aa aa aa aa 3’ 3’ 3’ aa 5’ 5’ 5’ tRNA Release factor N N Termination in E coli Yang Xu, College of Life Sciences
Q3 Initiation in Eukaryotes • Overview • Scanning • Initiation • Elongation and Termination Yang Xu, College of Life Sciences
Overview A comparison of the factors involved in prokaryotes and eukaryotes is given in Table 1. Yang Xu, College of Life Sciences
Bacteria Ribosome: 30S+50S →70S Few initiation factors (three factors) IF1, IF2, IF3 Elongation factors EF1A (EF-Tu), EF1B (EF-Ts), EF2 (EF-G) Release factors RF1, RF2, RF3 Ribosome recycling factor RRF mRNA is not capped Direct binding of 30S particle next to initiation codon (AUG) at Shine-Dalgarno sequence, 5’-AGGAGGU-3’ Translation coupled to transcription Eukaryotes Ribosome: 40S+60S → 80S Many initiation factors (14 factors) eIF1, eIF1A, eIF2, eIF2B, eIF3, eIF4A, eIF4B, eIF4E, eIF4F, eIF4G, eIF4H, eIF5, eIF5B, eIF6 Elongation factors eEF1, eEF2 Release factors eRF1 (or eRF2), eRF3 Most mRNA is capped at 5’ end and poly (A) at 3’ end 40S particle is recruited to 5’ cap structure or poly(A) tail or an internal ribosome entry site (IRES) Translation in always in cytoplasm apart from transcription Differences between prokaryotes and eukaryotes Yang Xu, College of Life Sciences
Scanning • Since there is no Shine-Dalgarno sequence in eukaryotic mRNA, the mechanism of selecting the start codon must be different. • Prof. Kozak proposed a scanning hypothesis in which the 40S subunit, already containing initiator tRNA, attaches to the 5’-end of the mRNA and scans along the mRNA until it finds an appropriate AUG. • Eukaryotic small ribosome submit complex bind to the 5’cap region of the mRNA and moves along it scanning for an AUG start codon. Yang Xu, College of Life Sciences
Initiation • Dissociation of ribosome • binding eIF1A (4C) and eIF3 to 40S • Recruit ternary complex • (fMet-tRNAi, eIF2, GTP) to 40S • Recruit mRNA to 43S → 46S particle • 46 S scans mRNA to reach AUG • Initiation factors leave and 80S joins • Elongation begins Yang Xu, College of Life Sciences
Initiation • Dissociation of ribosome binding of • eIF1A (eIF4C-A-site) and eIF3 to 40S • eIF3 is a huge complex (MW=0.69 MDa) of 11 subunits not homologous to IF3 • Possibly eIF5, eIF5B and eIF1 bind at this point as well • eIF1 seems to play a similar role as IF3-C • Recruit ternary complex (fMet-tRNAi, • eIF2, GTP) to 40S • eIF2 is a complex of three subunits (a, , ) of 123 kDa • GTP is loaded by eIF2B • Regulated by phosphorylation • Recruit mRNA to 43S • mRNA is bound to the eIF4F complex (4E, 4G, 4A) • Scan mRNA to reach AUG • Scanning requires a defined number of eIFs • Initiation factors leave and 80S joins • eIF5 is the exchange GEF for eIF2→ eIF2 release • Ribosome is GEF for eIF5B→ eIF5B release • Elongation begins Yang Xu, College of Life Sciences
Elongation and Termination Elongation • The protein synthesis elongation cycle in prokaryotes and eukaryotes is quite similar. • Three factors (EF-Tu, EF-Ts and EF-G) are required properties similar to their prokaryotic counterparts. • eEF1α, eEF1βγand eEF2 have the roles described for EF-Tu, EF-Ts and EF-G respectively. Termination • In eukaryotes, a single release factor, eRF, recognizes all three stop codons and performs the roles carried out by RF1 (or RF2) plus RF3 in prokaryotes. eRF requires GTP for activity, but it is not yet clear whether there is a eukaryotic equivalent of ribosome release factor required for dissociation of the subunits from the mRNA. Yang Xu, College of Life Sciences
Q 4 Translation Control and Post-translational Events • Translational control • Poly-proteins • Protein Targeting • Protein modification • Protein degradation Yang Xu, College of Life Sciences
Translational control In prokaryotes • Short antisense molecules can obscure ribosome binding; • The formation of stems and loops can inhibit exonuleases; • The relative stability to nucleases of parts of the polycistronic mRNA; • The binding of protein that prevent ribosome access. • In eukaryotes • Generally control protein amount by transcription of genes; • The repeats of the sequence 5’-AUUUA-3’ can make the mRNA for rapid degradation and thus limited translation; • Protein binding can mask the mRNA (masked mRNA) and prevent translation. Yang Xu, College of Life Sciences
Polyproteins • (1) Bacteriophage and viral transcipts and (2) many mRNAs for hormones in eukaryotes (e.g. pro-opiomelanocortin) are translated to give a single polypeptide chain that is cleaved subsequently by specific proteases to produce multiple mature protein from one translation product. The parent polypeptide is called polyprotein. Yang Xu, College of Life Sciences
Protein Targeting-I • Signal sequence • It is a short peptide sequences which composed of about 13~36 amino acids. • Function: The signal sequence causes the translating ribosome to bind factors that make the ribosome dock with a membrane and transfer the protein through the membrane as it is synthesized. • Usually the signal sequence is then cleaved off by signal peptidase. • Signal recognition particle (SRP) • SRP can recognize ribosomes with signal peptide of the nascent chain. • SRP receptor (docking protein) • SRP with the arrested ribosome can bind to SRP receptor on the cytosolic side of the Endoplasmic Reticulum (ER); • Ribosome receptor protein • When the ribosome becomes attached to ribosome receptor protein on the ER, SRP is released and can be re-used. • The ribosome is able to continue translation, and the nascent polypeptide chain is pushed through into the lumen of the ER. Yang Xu, College of Life Sciences
Protein Targeting-II • Glycosylation • The protein in ER is usually modified by glycosylation, and different patterns of Glycosylation seem to control the finial location of the protein. • Nuclear localization signal (nls) • Different N-terminal sequences can cause protein to be imported into mitochondria or chloroplasts, and the internal sequence –Lys-Lys-Lys-Arg-Lys, or any five continuous amino acids, can be a nuclear localization signal (nls). It cause the protein to be imported into the nucleus. Yang Xu, College of Life Sciences
The secretory pathwayin eukaryotes (co-translational targeting) Yang Xu, College of Life Sciences
Phosphorylation Protein modification The most common alterations to nascent polypeptides are those of cleavage and chemical modification. • Polypeptide cleavage (by amino- & carboxypeptideases) • To remove signal peptides; • To release mature fragments from polyprotein; • To remove internal peptides as well as • To trim both N- and C-termini. • Chemical modification: • Acetylation; • Hydroxylation; • Phosphorylation; • Methylation; • Glycosylation. Yang Xu, College of Life Sciences
26S protease complex Protein degradation A protein that is damaged, modified or has an inherently destabilizing N-terminal residue becomes ubiquitinylated by covalent linkage of molecules of the small, highly conserved, ubiquitin, via its C-terminal Gly, to lysine residues in the protein. The ubiquitinylated protein is digested by a 26S protease complex in a reaction that requires ATP and releases intact ubiquitin for re-use. In eukaryotes, N-terminal residue plays a critical role in inherent stability: • t1/2>20 hours: Ala, Cys, Gly, Met, Pro, Ser, Thr, Val (eight aa); • t1/2 2~30 min: Arg, His, Ile, Leu, Lys, Phe, Trp, Tyr (eight aa); • Destabilizing: Asn, Asp, Gln, Glu. (4aa) Yang Xu, College of Life Sciences
That’s all for Section Q Yang Xu, College of Life Sciences