Download

1 / 60
600 likes | 795 Views
Protein phosphorylation in modulation of cell functions. Jau-Song Yu ( 余兆松 ) Department of Cell and Molecular Biology, Institute of Basic Medical Sciences, Medical College of Chang Gung University. ( 長庚大學基礎醫學所分子生物學科 ). Reversible Phosphorylation of Proteins. OH. Protein/Enzyme.

E N D
Protein phosphorylation in modulation of cell functions Jau-Song Yu (余兆松) Department of Cell and Molecular Biology, Institute of Basic Medical Sciences, Medical College of Chang Gung University (長庚大學基礎醫學所分子生物學科)
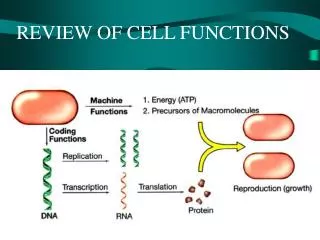
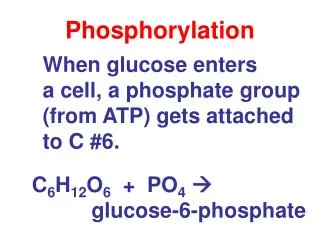
Reversible Phosphorylation of Proteins OH Protein/Enzyme Protein phosphatase Protein kinase Protein/Enzyme PO4 (Ser, Thr or Tyr) Cellular Processes: Metabolism, contractility, membrane transport and secretion, transcription and translation of genes, cell division, fertilization, memory, carcinogenesis, apoptosis, etc.
The 1992 Nobel Prize in Physiology or Medicine NOBELFÖRSAMLINGEN KAROLINSKA INSTITUTETTHE NOBEL ASSEMBLY AT THE KAROLINSKA INSTITUTE (12 October 1992) The Nobel Assembly at the Karolinska Institute has today decided to award the Nobel Prize in Physiology or Medicine for 1992 jointly to Edmond H. Fischer and Edwin G. Krebs for their discoveries concerning "reversible protein phosphorylation as a biological regulatory mechanism". Summary Thousands of proteins participate in a complex interplay in a cell. They are the tools of the living organism, regulating its reactions and activities. For example, proteins maintain the metabolic flux, dictate growth and cellular division, release hormones, and mediate muscular work. Protein interactions are strictly controlled. One of the most important regulatory mechanisms is reversible protein phosphorylation. This means that enzymes phosphorylate and dephosphorylate proteins. Both these enzymatic processes are in turn regulated, often in several steps, allowing amplification and fine control. The 1992 Nobel Prize in Physiology or Medicine is awarded to the American biochemists Edmond Fischer and Edwin Krebs. They purified and characterized the first enzyme of this type. Their fundamental finding initiated a research area which today is one of the most active and wide-ranging. Reversible protein phosphorylation is responsible for regulation of processes as diverse as mobilization of glucose from glycogen, prevention of transplant rejection by cyclosporin, and development of a cancer form like chronic myeloic leukemia.
Phosphoryl groups affect the structure and catalytic activity of proteins Glycogen phosphorylase (Glucose)n + Pi (glucose)n-1 + glucose 1-phosphate
Regulation of glycogen phosphorylase AMP Glucose PLP P-Ser14 Pyridoxal phosphate (PLP) Un-P: 20 aa (+) residues at its N terminus Interact with multiple acidic aa P-Ser14: interferes this interaction, more active conformation
The 2001 Nobel Prize in Physiology or Medicine 8 October 2001 The Nobel Assembly at Karolinska Institutet has today decided to award The Nobel Prize in Physiology or Medicine for 2001jointly to Leland H. Hartwell, R. Timothy (Tim) Hunt and Paul M. Nurse for their discoveries of "key regulators of the cell cycle" Summary All organisms consist of cells that multiply through cell division. An adult human being has approximately 100 000 billion cells, all originating from a single cell, the fertilized egg cell. In adults there is also an enormous number of continuously dividing cells replacing those dying. Before a cell can divide it has to grow in size, duplicate its chromosomes and separate the chromosomes for exact distribution between the two daughter cells. These different processes are coordinated in the cell cycle. This year's Nobel Laureates in Physiology or Medicine have made seminal discoveries concerning the control of the cell cycle. They have identified key molecules that regulate the cell cycle in all eukaryotic organisms, including yeasts, plants, animals and human. These fundamental discoveries have a great impact on all aspects of cell growth. Defects in cell cycle control may lead to the type of chromosome alterations seen in cancer cells. This may in the long term open new possibilities for cancer treatment.
SCIENCE, 298, 1912-34 (2002) The Protein Kinase Complement of the Human Genome G. Manning,1* D. B. Whyte,1 R. Martinez,1 T. Hunter,2 S. Sudarsanam1,3 We have catalogued the protein kinase complement of the human genome (the “kinome”) using public and proprietary genomic, complementary DNA, and expressed sequence tag (EST) sequences. This provides a starting point for comprehensive analysis of protein phosphorylation in normal and disease states, as well as a detailed view of the current state of human genome analysis through a focus on one large gene family. We identify518 putative protein kinase genes, of which 71 have not previously been reported or described as kinases, and we extend or correct the protein sequences of 56 more kinases. New genes include members of well-studied families as well as previously unidentified families, some of which are conserved in model organisms. Classification and comparison with model organism kinomes identified orthologous groups and highlighted expansions specific to human and other lineages. We also identified 106 protein kinase pseudogenes. Chromosomal mapping revealed several small clusters of kinase genes and revealed that 244 kinases map to disease loci or cancer amplicons. [public and Celera genomic databases, Incyte ESTs, in-house and GenBank cDNAs and ESTs) with a hidden Markov model (HMM) profile of the ePK domain]
Kinase distribution by major groups in human and model systems
Chromosomal Mapping and Disease Comparison of the kinase chromosomal map with known disease loci indicates that 164 kinases map to amplicons seen frequently in tumors (Knuutila et al., 1998) and 80 kinases map to loci implicated in other major diseases S. Knuutila et al., Am. J. Pathol. 152, 1107 (1998). DNA copy number amplications in human neoplasms: Review of comparative genomic hybridization studies.
Kinetic profiles of kinase activity and cell function change in cells Gene level Kinase A Kinase B Kinase c Time
Kinase assay in immunoprecipitate (IP) Cells *homogenization (10-cm dish/0.5 ml lysis buffer) *centrifugation (12000~15000 rpm, 15 min, 4oC) Supernatants *protein concentration determination *1 mg protein/0.5 ml extracts *add Ab against specific kinase (5 mg) *incubation (1 h, 4oC) *add protein A/G-S4B (50% v/v, 25 ml, shaking) *centrifugation (6000 rpm, 1min, 4oC) *wash/cfg 3 times in Buffer B Immunoprecipitates *suspended in 20 ml Buffer A *substrate (5-10 mg), [g-32P]ATP.Mg2+ (0.2-20 mM) *shaking for 10-30 min at RT *adding SDS-sample buffer SDS-PAGE Autoradiography cfg Lysis buffer-----10 mM Tris-HCl at pH 7.4, 2 mM EDTA, 1 mM EGTA, 1% Triton X-100, 1 mM benzamidine, 1 mM phenylmethylsulfonyl fluoride, 0.5 mg/ml aprotinin Buffer A --- 20 mM Tris-HCl at pH 7.0, 0.5 mM dithiothreitol Buffer B --- 0.5 M NaCl in buffer A (quantitative method)
JNK activity assay in IP (Chan et al., 2000)
Kinase assay by immunoblotting with phospho-specific Ab (Qualitative to semi-quantitative method)
Time post PDT (hr) C CL P 0 0.5 1 1.5 2 3 4 p-JNK2 p-JNK1 JNK1 JNK activity assay by Western blot (Hsieh et al., 2003)
Immunoblot analysis of auto-kinase during the activation and inactivation processes with a phospho-specific antibody against the identified phosphorylation-site sequence (Yu et al., Biochem. J. 1998)
Activation of kinases by osmotic shock (In-gel kinase assay) 37°C for 30 min/60 mg extracts 0.7 M NaCl at 37°C (Chan et al., 1999)
Elucidation of kinase pathway involved in specific signal events by specific activators and inhibitors
Activation of MAPK by osmotic shock and inhibition of osmotic shock-activated MAPK by PD98059 (+PD98059) (-PD98059) (-PD98059) (In-gel kinase assay) (Chan et al., 1999)
The specificities of protein kinase inhibitors: an update Bain J et al., Biochem J 371, 199-204 (2003)
PNAS USA 100, 15434–15439 (2003) Fig. 2. Optimization of adsorption and elution conditions for a functional p38 inhibitor matrix. Fig. 1. Identification of a p38 inhibitor analogue suitable for immobilization.
16-benzyldimethyl-n-hexadecylammonium chloride (16-BAC) Fig. 3. Efficient affinity purification of protein kinases specifically targeted by immobilized p38 inhibitor. HeLa whole cell lysate was subjected to PI 51 affinity chromatography, and the bound proteins were eluted with a combination of ATP and free PI 51.
Fig. 6. Structural determinants of SB 203580 sensitivity. Fig. 5. In vitro characterization of protein kinases inhibited by SB 203580.
Elucidation of kinase pathway involved in specific signal events by anti-sense oligonucleotide against specific kinase
Inhibition of JNK1 by anti-sense oligonucleotide attenuates caspase-3 activation by of photodynamic treatment JNK1 sense (5’-ATC ATG AGC AGA AGC AAG CGT GAC-3’) and anti-sense (5’-GTC ACG CTT GCT TCT GCT CAT GAT-3’) oligonucleotides (Chan et al., 2000)
Non-specific effects resulting from the introduction of siRNA appear to have three separate origins3: • 1. Lipid-mediated response4 • 2. Interferon response5 • 3. RISC-dependent off-target effects6-10 Dharmacon, having conducted considerable research on off-target effects, found that a combination of bioinformatics, chemical modifications, and siRNA pooling significantly reduce unintended targeting. These three innovations are included in the new ON-TARGETplus SMARTpool siRNA reagents. 3. Lin, X., et al. "siRNA-mediated off-target gene silencing triggered by a 7 nt complementation." Nucleic Acids Res. 33.14 (2005): 4527-35. 4. Fedorov, Y., et al. "Different delivery methods-different expression profiles." Nat Methods 2.4 (2005): 241. 5. Sledz, C., et al. "Activation of the interferon system by short-interfering RNAs." Nat Cell Biol 5.9 (2003): 834-9. 6. Semizarov, D., et al. "Specificity of short interfering RNA determined through gene expression signatures." PNAS 100.11 (2003): 6347-52. 7. Scacheri, P. C., et al. "Short interfering RNAs can induce unexpected and divergent changes in the levels of untargeted proteins in mammalian cells." PNAS 101.7 (2004): 1892-97. 8. Snove, O., and T. Holen. "Many commonly used siRNAs risk off-target activity." Biochem. Biophys. Res. Commun. 391.1 (2004): 256-63. 9. Jackson, A. L., et al. "Expression profiling reveals off-target gene regulation by RNAi." Nat. Biotech. 21.6 (2003): 635-7. 10. Jackson, A. L., and P. S. Linsley. "Noise amidst the silence: off-target effects of siRNAs?" Trends Genet. 20.11 (2004): 521-24.