Download

1 / 12
120 likes | 362 Views
The Rho GTPase activators CNF1 and DNT bacterial toxins have mucosal adjuvant properties Patrick Munro, et al. Presented by Jared Nelson. Introduction

E N D
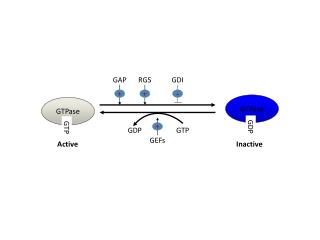
The Rho GTPase activators CNF1 and DNT bacterial toxins have mucosal adjuvant propertiesPatrick Munro, et al.Presented by Jared Nelson Introduction • There is, even today, a demand for more effective vaccines containing adjuvants that help stimulate an IgA(and IgG) response. Cholera toxin (CT) and other enterotoxins have been identified as potent IgA-stimulating adjuvants that could be used in vaccines. • Cytotoxic Necrotizing Factor 1(CNF1), found in uropathogenic E. coli, is structurally and functionally related to E. coli CNF2, Yersinia pseudotuberculosis CNFy, and Bordetella Dermonecrotic toxin(DNT). CNF1 is an A-B toxin. CNF1 and DNT both use similar mechanisms to invade and intoxicate host cells and to allow bacterial cells to invade and persist in host cells.
CNF1 and DNT are both taken up by host cells via their amino-terminal domains into vesicles. Their translocation domains are used in acidic pHs to eject the toxins out of their vesicles into the cytoplasm. After this, these toxins convert Rho proteins into their active forms by stopping their GTPase functioning via deamidation or transglutamination. Then ubiquitin regulates the digestion of these Rho proteins by proteases/proteasomes. • The activation of Rho proteins, such as Rac and Cdc42 (particularly via CNF1), is all that is needed to send a signal cascade that leads to generating IL-8, MCP-1, and MIP-3-alpha (cytokines that are involved in inflammation and directly or indirectly in the regulation of the adaptive immune response). • This experiment points to CNF1 and DNT as being agents used to elicit a strong T-cell mediated and B-cell mediated immune reaction with a hydrophilic protein antigen named ovalbumin (OVA). They claim that Rho proteins(GTPases), when activated by CNF1 and DNT, are directly responsible for the regulation and activation of the above adaptive immunity.
Materials and Methods • I. CNF1 and CNF1-C866S(mutant inactivated form of CNF1) toxins were made and purified. CNF1 samples were subjected to a Hep2 multinucleation assay to determine their activity. CNF1-CD and DNT-CD (CD stands for ‘catalytic domain’) were also made and purified. Rac (a Rho GTPase) activating and digestion by DNT-CD was observed with the “GST protein pull-down” experiment. • II. Each lab mouse was given (orally) one of the following toxins: CNF1, CNF1C866S, CNF1-CD, DNT-CD, or CT along with OVA, except for the OVA-only controls. Mice were given this cocktail 2 or 3 times, with 10-12 days between times. • III. Anti-OVA antibodies were detected by “solid phase ELISA” using HRP-tagged goat anti-mouse antibodies.
IV. Anti-OVA IgA and IgG were extracted from tissue samples from mice. Supernatants from these tissue samples and/or sera were analyzed via ELISA. • V. Mice were given CNF1 or CNF1-C866S, and small intestines were processed and sectioned using paraffin( The toxins were supposedly given without OVA, perhaps to confirm that only anti-OVA antibodies were produced).
Results • Mice given OVA and 10 ug CNF1 produced a very high titer of anti-OVA IgG. When the mice were given cholera toxin, the anti-OVA IgG titer was almost as high as the above result. • IgGs specific for CNF1 were not found. • NOTE: Only anti-OVA antibodies were detected. • The anti-OVA IgG titer yielded by 1 ug CNF1 was only slightly greater than that produced by OVA alone (the + control). These titers were several times less than the CT and CNF1 results. • When mice were given 10 ug of CNF1-C866S mutant ( - control), the IgG titer was next to zero. • Therefore, only a high dose of active CNF1 will give a high IgG titer.
In figure 2A(GST protein pull-down), DNT temporarily activated Rac, since Rac is digested afterwards. The RacGTP band is getting larger overall as time goes on simply because the GTP is not being hydrolyzed = no GTPase activity in Rac. Then notice the last RacGTP band is smaller compared to the next to last one, probably because Rac is being digested. The last lane, showing actin, apparently represents actin rearrangement, although it doesn’t appear to prove this. • Contrary to what the diagram tells us verbatim, fig. 2B shows how effectively CNF1-CD and DNT-CD intoxicate (activate Rac within ) host cells(intestinal endothelial). DNT is clearly much more effective than CNF1 in terms of its intoxicating effect and in terms of the IgG titer DNT stimulates (fig 2C).
Fig 3 shows no measureable levels of CNF1-C866S-stimulated IgA nor IgG, except for a slight amount of IgG1. • Compare this to varying but similar levels of IgG subclasses, such as IgG2a, and IgA, produced from CNF1 and CT stimulation. The amounts of IgA were low in both of these conditions; the amounts of all the varieties of IgGs were more or less significantly higher than those of IgA.
When mice were given OVA and CNF1 or CNF1C866S(10 ug for either one), the CNF1 produced IgA titer was several times greater than that produced by CNF1C866S or in an OVA-only control.
In fig. 5, Intestinal tissue preparations (with paraffin cross-sections) of CNF1 or CNF-C866S-treated mice were not very different from those of a bicarbonate treated control. I’m not sure why this is, since I would expect to see signs of inflammation, at least.
Discussion and Conclusions • CNF1 boosts IgA and IgG-based adaptive immunity with respect to a protein antigen, OVA. CNF1 is an adjuvant because of its (at least) deamidase activity. CT has adjuvant effects that are close, in terms of what degree of titer of which antibodies, whether IgG or IgA, are produced, to those of CNF1. All antibodies produced were specific to OVA. • CNF1-C866S, the mutant, doesn’t influence the adaptive immune reactions very much at all. • Both CNF1 and DNT had a temporary activating effect on Rac in relating ways; therefore, DNT has the same kinds of adjuvant effects, in which the active Rho proteins like Rac are directly responsible for the adaptive immune effects.
Future questions • They haven’t drawn any conclusions about whether 1. the cytokine MIP3-alpha activation/regulation of IgA adaptive immunity and 2. an active Cdc42 influencing the process of dendritic cell maturing are important parts of CNF1’s adaptive immunity boosting influence. • Could CNF1’s toxic effect as explained in this study impart any benefit to E. coli? The only benefit I can think of is one that these researchers mentioned about active CNF1 rearranging the host cell cytoskeleton, allowing the E. coli to invade the cells and persist there through resistance to signals that lead to host cell apoptosis. • How else could we target or influence Rho proteins to regulate/boost the adaptive immune activity?