Download

1 / 34
350 likes | 548 Views
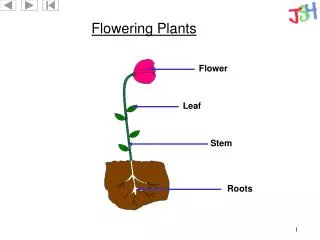
Flowering. Cosa induce la fioritura Come avviene la transizione da stadio vegetativo a stadio fiorale Il Meristema Apicale Vegetativo Il Meristema Apicale Fiorale Il differenziamento degli organi fiorali Ruolo dei geni omeotici: il modello ABC. TRANSITION TO FLOWERING.

E N D
Flowering • Cosa induce la fioritura • Come avviene la transizione da stadio vegetativo a stadio fiorale • Il Meristema Apicale Vegetativo • Il Meristema Apicale Fiorale • Il differenziamento degli organi fiorali • Ruolo dei geni omeotici: il modello ABC
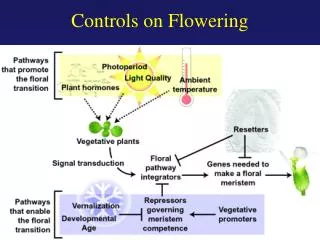
TRANSITION TO FLOWERING 4 pathways regolano questo processo 1) Vernalizzazione 2) Fotoperiodo 3) GA 4) Autonomo 1-2 regolano il flowering in risposta a segnali ambientali 3-4 agiscono in modo indipendente dagli stimoli ambientali
Arabidopsis è una long-day facoltativa • Il pathway di fotoperiodo induce fioritura in risposta all’aumento del rapporto giorno/notte • Il pathway di vernalizzazione rende Arabidopsis competente alla fioritura dopo esposizione a basse temperature per un lungo periodo • Il pathway autonomo reprime la fioritura costitutivamente • Il pathway delle gibberelline induce la fioritura e le GAs sono necessarie in condizioni di fotoperiodo non-inducing • C’è variazione nei tempi di fioritura fra ecotipi di Arabidopsis: late-flowering senza vernalizzazione v/s gli early-flowering, si differenziano per variazione allelica nei loci FRI e FLC. I primi hanno alleli FRI e FLC dominanti, gli altri hanno alleli fri e flc non funzionali o deboli
VRN2 VRN1 metilazione/remodeling cromatina Pathway Autonomo FPA FVE FY FCA LD FLD FRI FLC azioneGA Saccarosio LFY Fase Vegetativa Fioritura Vernalizzazione
Vernalizzazione Complesso PAF1 Pathway autonomo Fotoperiodo FLC SOC1 Gibberelline LEAFY AP1 Transizione fiorale FT
FT: • piccola proteina globulare di 20 kDa, espressa nei tessutii floematici di cotiledoni e foglie • è un target diretto di CO, regolatore trascrizionale del pathway di fotoperiodo • il ruolo di FT come promotore del flowering è conservato in Arabidopsis e riso • è a valle di FLC e, in particolare, del pathway di fotoperiodo di induzione della fioritura • la sua attività è regolata da FD, una proteina bZip • FT e FD sono sufficienti ad attivare l’espressione dei geni marcatori fiorali (floral meristem identity genes) • Come FT regola l’attività di FD?
FD: • identificata mediante “two-hybrid”, come “interactor” di FT • FD è necessario per l’attività di FD • FT e FD sono sufficienti per attivare l’espressione dei geni marker del flowering • FT agisce nel nucleo in un complesso contenente FD direttamente sui promotori dei geni target (chIP) • E’ noto da tempo che il fotoperiodo è percepito nelle foglie, che rilasciano un segnale sistemico, il così detto “florigeno”. • FT è espresso, tramite CO, nel floema fogliare, ma la sua attività nel complesso FT-FD avviene nel meristema apicale • L’ipotesi più semplice è che l’induzione di FT nel sistema vascolare fogliare porti alla presenza della proteina FT nel meristema apicale • Alternativamente, l’espressione di FT nelle foglie può attivare un “florigeno” che induce l’espressione di FT nel meristema
Transizione da stadio vegetativo a stadio fiorale regolata da fattori ambientali e di sviluppo • Vernalizzazione: acquisizione della capacità alla fioritura in seguito a trattamento a freddo • il segnale di vernalizzazione deve essere trasmesso attraverso diversi cicli di divisione cellulare, NON è però trasmesso alla progenie • FLC, FRI:repressori di fioritura • FRIagisce interagendo con FLC • FLCregolato sia a livello trascrizionale che post-trascrizionale, rappresenta il segnale che è inibito dalla vernalizzazione
Vernalizzazione: • non è graft-trasmissibile • effetti mitoticamente trasmissbili, NON trasmessi alla meiosi • VRN1/2:responsabili del mantenimento dello stato di vernalizzazione • VRN1codifica per una Myb-related DNA binding protein • VRN2codifica per una proteina di tipo Polycomb (Enhancer of zeste-like), zinc-finger, localizzata nel nucleo, necessaria per la stabilizzazione della repressione di FLC • Il complesso contenente VRN1/2 ha attività dimetilasi istonica
Vernalizzazione supera l’effetto di FRI, reprimendo l’espressione di FLC repressione epigenetica di FLC • FLCè espresso principalmente in zone mitoticamente attive Memoria Cellulare della Vernalizzazione dovuta a struttura cromatinica alterata in FLC • La recente caratterizzazione di regolatori positivi e negativi di FLC ha mostrato che alcuni di loro sono coinvolti nella modificazione cromatinica di FLC. Queste modificazioni cromatiniche rappresentano un livello ulteriore di regolazione dell’expressione genica
VRN1 e VRN2 hanno profili di espressione più “ampi” di FLC e NON sono regolati dalla vernalizzazione • come possono reprimere FLC solo in seguito a vernalizzazione? • VIN3(Vernalization insensitive 3) • è indotto da freddo e di conseguenza FLC viene represso • VIN3 codifica per una HD protein. • Insieme a VRN1 (Myb TF) e a VRN2 (EnZeste) puo’ partecipare al remodeling di FLC
Modello di Silencing di FLC mediato da vernalizzazione FLC attivo:H3 acetilato FLC inattivo:H3 deacetilato il complesso HDAC/VIN3 lega H3 deacetilato il complesso VRN1/VRN2 metila H3 in Lys 9 e 27 il complesso VRN1/VRN2 lega stabilmente H3, mantenendo lo stato di repressione HP1 (Heterocromatin Protein1) lega H3 deacetilato, contribuendo al silenziamento di FLC
Ac Ac Ac Ac Ac Ac H3 H3 H3 H3 H3 H3 FLC winter Inactive FLC HDAC VIN3 HDAC VIN3 HDAC VIN3 winter Active H3 HDAC VIN3 Histone deacetylase complex Histone H3
H3 H3 H3 FLC HP1 HDAC VIN3 HDAC VIN3 HDAC VIN3 spring HP1 HP1 MEK9/27 MEK9/27 MEK9/27 MEK9/27 MEK9/27 MEK9/27 H3 H3 H3 FLC Ez VRN1/2 Ez VRN1/2 Ez VRN1/2 Ez VRN1/2 Ez VRN1/2 Ez VRN1/2 Ez VRN1/2 Inactive Stable repression Heterocromatin Protein1 Lega H3 dimetilato Enhancer of zeste-like VRN1/VRN2 HP1
Attivazione dell’espressione di FLC La trimetilazione dell’istone H3 su Lys4 (H3-K4) è associata con la cromatina di geni attivi La metilazione di H3-K4 avviene a 3 livelli: mono, di e trimetil In lievito e Drosophila, la presenza di un K4 trimetilato definisce uno stato attivo di espressione genica Come???? In lievito, l’ RNAPol Associated Factor1 (PAF1) si associa con il complesso RNA polimerasi II durante la trascrizione. Il suo ruolo è di recrutare SET1, una H3-K4 metiltrasferasi, sui geni target. SET1 determina la trimetilazione H3-K4 principalmente nella porzione 5’ delle regioni attivamente trascritte.
Mostrano un aumentato livello di trimetilazione H3-K4 di FLC: • I mutanti late-flowering del pathway autonomo; • Le linee late-flowering che hanno un locus FRI forte • Quindi; • FRI promuove la trimetilazione H3-K4 della regione cromatinica di FLC, aumentando il livello di espressione di FLC • I loci del pathway autonomo sono repressori di questa modificazione cromatinica In Arabidopsis ci sono 5 geni “parenti” di FLC: FLM, MAF1, MAF3, MAF4 e MAF5 Come FLC, FLM e MAF2 sono repressori della fioritura ma, a differenza di FLC, reprimono la fioritura in condizioni di fotoperiodo non-inducing
In lievito, esiste un solo gene per SET1, mentre Arabidopsis ha diverse potenziali H3-K4 trimetiletransferasi, basati su omologia di sequenza del dominio SET. • In particolare, mutazioni in Early Flowering in Short Days (EFS) comporta: • riduzione del livello di trimetilazione H3-K4 di FLC, • soppressione del late-flowering mediato da FRI • soppressione dell’espressione di FLC • EFSrecrutato da PAF1 per metilare i geni targets???
In lievito ISW1p, proteina ATPasi chromatin-remodeling, preferenzialmente lega H3-K4 di e trimetilati e la metilazione cromatinica H3-K4 di certi geni attivamente trascritti è richiesta per l’associazione di ISW1p in vivo. Un omologo di ISW1p di Arabidopsis, PIE1, è necessario per l’espressione di FLC. PIE1 lega K4 trimetilate e rimodella la cromatina di FLC e dei suoi “parenti” per aumentare l’espressione genica. Lievito ed Arabidopsis usano la trimetilazione H3-K4 in modo molto simile
PolII Il complesso PAF1 include: ELF7, ELF8 e VIP4 E’ associato a PolII PolII EFS PolII EFS PIE1 PIE1 PolII EFS PIE1 PIE1 FLCmRNA LOCUS FLC Senza vernalizzazione EFS H3-K4 metiltransferasi PIE1 Lega K4 trimetilato e causa remodeling ulteriore della cromatina di FLC e aumento della tracsrizione
LOCUS FLC dopo vernalizzazione Stato eterocromatinico No legame di PAF1
Flowering • Cosa induce la fioritura • Come avviene la transizione da st. vegetativo a st. fiorale • Il Meristema Apicale Vegetativo • Il Meristema Apicale Fiorale • Il differenziamento degli organi fiorali • Ruolo dei geni omeotici: il modello ABC
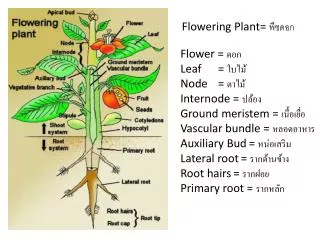
Shoot Apical Meristem (SAM) • Ha 3 diverse attività: • automantenimento • produzione di foglie • formazione dello stelo • Diviso in 3 zone: • CZ (Central Zone)all’apice del meristema, scarsa attività di divisione cellulare,automantenimento • PZ (Peripheral Zone), circonda la central zone, % divisione cellulare alta,produzione di foglie • Rib (Rib Zone), dietro la central zone, % divisione cellulare alta,formazione dello stelo
Differenziamentocome risultato del controllo didivisione cellulare(controllo dei piani di divisione) eallungamento/espansione cellulare • 3 strati clonali distinti nel SAM, caratterizzati da diversimarcatori molecolari • L1distinto già durante l’embriogenesi, pattern di divisioni cellulari anticlinali • L2pattern di divisioni cellulari anticlinali • L3divisioni cellulari secondo piani diversi, approvvigionamento delle cellule centrali
Arabidopsis Shoot Apical Meristem (SAM) • durante la fase vegetativa, contribuisce allo sviluppo dello stelo e dei primordi fogliari • durante la fase riproduttiva produce meristemi fiorali • Due classi di geni hanno un ruolo nel determinare i meristemi apicali e fiorali: • Geni Meristem-Structure: • STM (TF), CLV1/3 (recep. kinase) • Geni Meristem-Identity
SAM Strati clonali Floral Meristem L1 L2 L1, L2 e L3 partecipano tutte all’organogenesi L1: epidermide di fusto, foglie, fiori L2: mesodermide e cellule germinali L3: tessuto vascolare L3 Zone istologiche CZ PZ PZ RZ SAM
Meristemi apicale e fiorale sono“indeterminati”in quanto mantengono la loro attività per un tempo indefinito Geni regolativi Controllano il destino/identità cellulare nel meristema fiorale. Nello sviluppo del fiore, il primo segno di differenziamento è l’attivazione dei geni MADS per l’identità degli organi MADS family of transcription factors: MCM1 (yeast), AGAMOUS e DEFICIENS (Arabidopsis), SERUM Response Factor (human)
DNA binding K MADS C I dimerizzazione multimerizzazione K keratin-like I intervening C C-terminal MADS-box Genes • I geni MADS sono dominanti non solo fra gli Organ Identity Genes ma sono anche presenti come Meristem Identity Genes, Intemediate genes, Cadastral genes ed anche come targets più a valle
Cadastral Genes Fattori Ambientali Late and Early Flowering Genes FCA LD CO AP2 Meristem Identity Genes SUP TFL1 AP2 LFY CAL AP1 BEL1 SIN1 ELF1 LUG Intermediate Genes UFO AGL2 AGL9 AGL4 AG Organ Identity Genes AP2 PI AG AGL11 AP3 AP1 A D B C GENI DOWNSTREAM sconosciuti TFs MADS
Organ Identity Genes Geni omeotici geni A: determinano i sepali geni A + B: determinano i petali geni B + C: determinano gli stami geni C: determinano i carpelli • Appartengono alla famiglia dei geniMADS • EspressiSOLOnelle regioni del fiore in sviluppo dove è necessaria la loro attività • Come sono attivati i geni omeotici fiorali?? • Come è regolata la loro espressione in regioni cosi delimitate spazialmente?
Il modello ABC Organi fiorali Organ identity Genes LFY, TF Zn finger
SUP LUG AP2 AG A AP1 AP2 B AP3 PI C AG 2) Cadastral Genes 1) Organ Identity Genes SUP LUG AP2 AG 3) Floral Meristem Identity Genes • Ogni verticillo (whorl) contiene espressione unica di 1/2 gruppi di attività dei geni 1 • Le attività A e C si reprimono mutualmente, i.e. almeno alcuni dei geni delle attività A e C sono sia 1 (organ identity) che 2 (cadastral)
Studi di genetica reverse sui geni MADS hanno evidenziato l’esistenza di altri 2 gruppi: D ed E I geni con funzione D avrebbero un ruolo nello sviluppo dell’ovulo I geni con funzione E avrebbero un ruolo di co-fattori che lavorano con i geni ABC nei 3 verticilli interni b) I geni MADS rappresentativi di ogni gruppo degli organ identity genes. Questi loci sono membri della sottofamiglia MIKC di MADS
1) Meristem Identity Genes Necessari per l’induzione dei geni della classe 3, sono regolatori positivi, i.e. la loro assenza comporta la conversione dei fiori in germogli 2) Cadastral Genes Regolatori spaziali della classe 1, delimitando le “boundaries” di espressione, prevenendo quindi espressione ectopica di 3 3) Organ Identity Genes Corrispondono ai geni omeotici selector di Drosophila. Controllano “organ identity” mediante attivazione dei geni downstream
Ha un ruolo precoce nel promuovere l’attività dei Meristem Identity Genes Fiorali LEAFY (LFY) • In seguito è necessario per la normale espressione dei geni ABC Queste due funzioni possono essere separate • Il mutante lfy ha fiori senza petali ne’ stami, e presenta organi foglie, sepali e carpelli-like • Ha un ruolo fondamentale nell’induzione degli “Organ Identity Genes” di classe B • L’ antigene LFY è stato individuato in cellule in cui il suo mRNA non è presente • La proteina LFY è attiva in queste cellule: attività “long range” di un TF esportato • La proteina LFY lega DNA in vitro, e funziona come attivatore trascrizionale