Download

1 / 114
1.28k likes | 1.83k Views
Plant. You’ll leaf this knowing more!. Form and Function. A presentation of Chapters 35 – 39 by Clarence Lewis, Megan Baxter, Josh Newton, and Vasu Patel. Chapter. 35. Plant Structure, Growth, and Development. Reproductive shoot (flower). Terminal bud. Node. Internode. Terminal bud.

E N D
Plant You’ll leaf this knowing more! Form and Function A presentation of Chapters 35 – 39 by Clarence Lewis, Megan Baxter, Josh Newton, and Vasu Patel
Chapter 35 Plant Structure, Growth, and Development
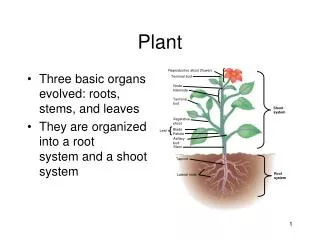
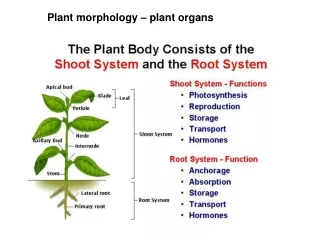
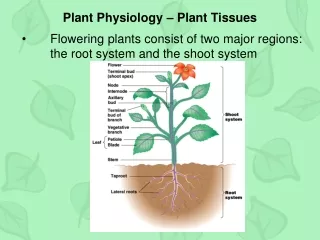
Reproductive shoot (flower) Terminal bud Node Internode Terminal bud Shoot system Vegetative shoot Blade Petiole Leaf Axillary bud Stem Taproot Root system Lateral roots Basic organs ∙Morphology is an organism’s external form. ∙Vascular plants have three basic organs: roots, stems, and leaves. ∙A root is an organ that anchors a vascular plant in the soil, absorbs minerals and water, and stores food. ∙A stem is an organ consisting of alternating nodes, the points at which leaves are attached, and internodes, the stem segments between nodes. ∙Leaves are the main photosynthetic organs of most plants, although green stems are also photosynthetic. Figure 35.2 An overview of a flowering plant.
Dermal tissue Ground tissue Vascular tissue Tissue systems ∙Plants are composed of three tissue systems: dermal, vascular, and ground. ∙The dermal tissue is the outer covering. ∙Vascular tissue, continuous throughout the plant, is involved in the transport of materials between roots and shoots. ∙The functions of ground tissue include photosynthesis, storage, and support. Figure 35.8 The three tissue systems.
Cell types ∙Plant tissues are composed of three basic cell types: parenchyma, collenchyma, and sclerenchyma. ∙Parenchyma cells perform most of the metabolic functions of the plant, synthesizing and storing various organic products. ∙Collenchyma cells have thicker primary walls than parenchyma cells, though the walls are unevenly thickened. ∙Sclerenchyma cells have thick secondary walls usually strengthened by lignin and function as supporting elements of the plant. ∙Xylem is vascular plant tissue consisting mainly of tubular dead cells that conduct most of the water and minerals upward from the roots to the rest of the plant. ∙Phloem is vascular plant tissue consisting of living cells arranged and elongated tubes that transport sugar and other organic nutrients throughout the plant.
Plant types ∙A major difference between plants and most animals is that plant growth is not limited to an embryonic period. ∙Annual plants complete their life cycle—from germination through flowering and seed production to death—in a single year or less. ∙The life of a biennial plant spans two years. ∙Often, there is an intervening cold period between the vegetative growth season and the flowering season. ∙Plants such as trees, shrubs, and some grasses that live many years are perennials. ∙Perennials do not usually die from old age, but from an infection or some environmental trauma.
Indeterminate growth ∙A plant is capable of indeterminate growth because it has perpetually embryonic tissues, called meristems, in its regions of growth. ∙The pattern of plant growth depends on the location of meristems. ∙Apical meristems, located at the tips of roots and in the buds of shoots, supply cells for the plant to grow in length. ∙Each spring and summer, as the primary growth extends the shoot, secondary growth thickens the parts of the shoot that formed in previous years.
Guard cells Key to labels Figure 35.18 Leaf anatomy. Dermal Stomatal pore Ground Vascular Epidermal cell Sclerenchyma fibers 50 µm Cuticle (b) Surface view of a spiderwort (Tradescantia) leaf (LM) Stoma Upper epidermis Palisade mesophyll Bundle- sheath cell Spongy mesophyll Lower epidermis Guard cells Cuticle Vein Xylem Vein Air spaces Guard cells Guard cells Phloem 100 µm Transverse section of a lilac (Syringa) leaf (LM) (c) (a) Cutaway drawing of leaf tissues
Revolution ∙Molecular biology is revolutionizing the study of plants. ∙Much of this research has focused on Arabidopsis thaliana, a small weed in the mustard family.
Growth ∙Growth involves both cell division and cell expansion. ∙Cell division in meristems increases cell number, increasing the potential for growth. ∙The plane (direction) and symmetry of cell division are important determinants of plant form. ∙While mitosis results in symmetrical redistribution of chromosomes between daughter cells, cytokinesis may be asymmetrical. ∙The plane in which a cell will divide is determined during late interphase. ∙The greatest expansion of a plant cell is usually oriented along the plant’s main axis.
Morphogenesis ∙Morphogenesis depends on pattern formation. ∙Morphogenesis organizes dividing and expanding cells into multicellular tissues and organs. ∙Developmental biologists are accumulating evidence that gradients specific molecules, generally proteins or mRNAs, that provide positional information. ∙One type of positional information is polarity, the identification of the root end and shoot end along a well-developed axis. ∙Other genes that regulate pattern formation and morphogenesis include the homeotic genes, which mediate many developmental events, such as organ initiation.
Cell location ∙Clonal analysis of the shoot apex emphasizes the importance of a cell’s location in its developmental fate. ∙In the process of shaping a rudimentary organ, patterns of cell division and cell expansion affect the differentiation of cells by placing them in specific locations relative to other cells. ∙One approach to studying the relationship among these processes is clonal analysis, mapping the cell lineages (clones) derived from each cell in an apical meristem as organs develop. ∙In plants, a cell’s developmental fate is determined not by its membership in a particular lineage but by its final position in an emerging organ.
Floral phase ∙Genes controlling transcription play key roles in a meristem’s change from a vegetative to a floral phase. ∙Unlike vegetative growth, which is indeterminate, the production of a flower by an apical meristem terminates primary growth of that shoot tip as the apical meristem develops into the flower’s organs. ∙Once a shoot meristem is induced to flower, positional information commits each primordium arising from the flanks of the shoot tip to develop into a specific flower organ. ∙Organ identity genes code for transcription factors. ∙The ABC model of flower formation identifies how these genes direct the formation of four types of floral organs.
Sepals Figure 35.34 The ABC hypothesis. Petals (a) A schematic diagram of the ABChypothesis. Studies of plant mutationsreveal that three classes of organ identitygenes are responsible for the spatial patternof floral parts. These genes are designated A,B, and C in this schematic diagram of a floralmeristem in transverse view. These genesregulate expression of other genesresponsible for development of sepals,petals, stamens, and carpels. Sepals developfrom the meristematic region where only Agenes are active. Petals develop where bothA and B genes are expressed. Stamens arisewhere B and C genes are active. Carpels arisewhere only C genes are expressed. Stamens Carpels A B C C gene activity B + C gene activity A + B gene activity A gene activity B B Active genes: B B B A A A B A B B C A C C A A C C C C A A A A C A C C C C C C C C A B B A A B B A Whorls: Carpel Stamen Petal Sepal Wild type Mutant lacking A Mutant lacking B Mutant lacking C (b) Side view of organ identity mutant flowers. The phenotype of mutants lacking a functional A, B, or C organ identify gene can be explained by combining the model in part (a) with the rule that is A or C activity is missing, the other activity occurs through all four whorls.
Chapter 36 Resource Acquisition and Transport in Vascular Plants
Selective permeability ∙Transport at the cellular level depends on the selective permeability of membranes. ∙The selective permeability of a plant cell’s plasma membrane controls the movement of solutes between the cell and the extracellular solution. ∙Transport proteins embedded in the membrane can speed movement across the membrane. Others act as selective channels, providing a selective passageway across the membrane. For example, the membranes of most plant cells have potassium channels that allow potassium ions (K+) to pass, but not similar ions, such as sodium (Na+). ∙Phylloxtaxy is the arrangement of leaves on the shoot of a plant. ∙Mycorrhizae is a mutalistic association of plan roots and fungus.
CO2 O2 Light 1 2 3 4 Through stomata, leaves take in CO2 and expel O2. The CO2 provides carbon for photosynthesis. Some O2produced by photosynthesis is used in cellular respiration. H2O Sugar Sugars are produced by photosynthesis in the leaves. Sugars are transported as phloem sap to roots and other parts of the plant. Transpiration, the loss of water from leaves (mostly through stomata), creates a force within leaves that pulls xylem sap upward. 6 5 7 Water and minerals are transported upward from roots to shoots as xylem sap. O2 Roots absorb water and dissolved minerals from the soil. H2O Roots exchange gases with the air spaces of soil, taking in O2 and discharging CO2. In cellular respiration, O2 supports the breakdown of sugars. CO2 Minerals Figure 36.2 An overview of resource acquisition and transport in a vascular plant.
Figure 36.3 Proton pumps provide energy for solute transport. EXTRACELLULAR FLUID CYTOPLASM – + H+ + – ATP H+ – + H+ Proton pump generates membrane potential and H+ gradient. H+ H+ H+ – H+ + H+ – + Proton pumps ∙Proton pumps play a central role in transport across plant membranes. ∙The most important active transport protein in the plasma membrane of plant cells is the proton pump. ∙The proton gradient also functions in co-transport, in which the downhill passage of one solute (H+) is coupled with the uphill passage of another, such as NO3- or sucrose. ∙The role of proton pumps in transport is a specific application of the general mechanism called chemiosmosis, a unifying process in cellular energetics.
Water potential ∙Differences in water potential drive water transport in plant cells. ∙In the case of a plant cell, the direction of water movement depends on solute concentration and physical pressure. ∙Plant biologists measure psi in units called megapascals (MPa), where one MPa is equal to about 10 atmospheres of pressure. ∙The combined effects of pressure and solute concentrations on water potential are incorporated into the following equation, where psip is the pressure potential and psis is the solute potential (or osmotic potential). psi = psip + psis ∙A walled cell with a greater solute concentration than its surroundings will be turgid, or firm. ∙Turgor pressure is the force directed against a plant cell wall after the influx of water and swelling of the cell due to osmosis.
Compartmental structure ∙Vacuolated plant cells have three major compartments. ∙While the thick cell wall helps maintain cell shape, it is the cell membrane, not the cell wall, which regulates the traffic of material into and out of the protoplast. ∙The membrane that bounds the vacuole, the tonoplast, regulates molecular traffic between the cytosol and the contents of the vacuole, called the cell sap. ∙In most plant tissues, two of the three cellular compartments are continuous from cell to cell.
Cell wall Transport proteins in the plasma membrane regulate traffic of molecules between the cytosol and the cell wall. Transport proteins in the vacuolar membrane regulate traffic of molecules between the cytosol and the vacuole. Cytosol Figure 36.11 Cell compartments and routes for short-distance transport. Vacuole Key Vacuolar membrane (tonoplast) Symplast Plasmodesma Plasma membrane Apoplast Transmembrane route Apoplast The symplast is the continuum of cytosol connected by plasmodesmata. The apoplast is the continuum of cell walls and extracellular spaces. Symplast Symplastic route Apoplastic route (b) Transport routes between cells. At the tissue level, there are three passages: the transmembrane, symplastic, and apoplastic routes. Substances may transfer from one route to another. (a) Cell compartments. The cell wall, cytosol, and vacuole are the three main compartments of most mature plant cells.
Mineral absorption ∙Root hairs, mycorrhizae, and a large surface area of cortical cells enhance water and mineral absorption. ∙Much of the absorption of water and minerals occurs near root tips, where the epidermis is permeable to water and where root hairs are located. ∙As the soil solution moves along the apoplast into the roots, cells of the epidermis and cortex take up water and certain solutes into the symplast. ∙Most plants form partnerships with symbiotic fungi to absorb water and minerals from soil.
Mineral pathway ∙The endodermis functions as a selective sentry between the root cortex and vascular tissue. ∙Water and minerals in the root cortex cannot be transported to the rest of the plant until they enter the xylem of the vascular cylinder. ∙Those minerals that reach the endodermis via the apoplast are blocked by the Casparian strip in the walls of each endodermal cell. ∙To enter the vascular cylinder, minerals must cross the plasma membrane of the endodermal cell and enter the vascular cylinder via the symplast. ∙The last segment in the soil-to-xylem pathway is the passage of water and minerals into the tracheids and vessel elements of the xylem.
Xylem sap Outside air Y = –100.0 MPa Mesophyll cells Stoma Water molecule Leaf Y (air spaces) = –7.0 MPa Transpiration Atmosphere Leaf Y (cell walls) = –1.0 MPa Xylem cells Adhesion Cell wall Water potential gradient Cohesion, by hydrogen bonding Trunk xylem Y = – 0.8 MPa Cohesion and adhesion in the xylem Water molecule Root xylem Y = – 0.6 MPa Root hair Soil Y = – 0.3 MPa Soil particle Water Water uptake from soil Figure 36.15 Ascent of xylem sap. Xylem sap ∙The ascent of xylem sap depends mainly on transpiration and the physical properties of water. ∙Root pressure causes guttation, the exudation of water droplets that can be seen in the morning on the tips of grass blades or the leaf margins of some small, herbaceous dicots. ∙The mechanism of transpiration depends on the generation of negative pressure (tension) in the leaf due to the unique physical properties of water ∙The water film at the surface of leaf cells has a negative pressure, a pressure less than atmospheric pressure.
Light absorption ∙The tension generated by adhesion and surface tension lowers the water potential, drawing water from an area of high water potential to an area of lower water potential. ∙These features are morphological adaptations to enhance the absorption of light for photosynthesis. ∙To make food, a plant must spread its leaves to the sun and obtain CO2 from air. ∙Transpiration also results in evaporative cooling, which can lower the temperature of a leaf by as much as 10–15°C relative to the surrounding air. ∙The stomatal density of a leaf is under both genetic and environmental control.
Sugar ∙Phloem translocates its sap from sugar sources to sugar sinks. ∙In contrast to the unidirectional flow of xylem sap from roots to leaves, the direction that phloem sap travels can vary. ∙A storage organ, such as a tuber or a bulb, may be either a source or a sink, depending on the season. ∙A sugar sink usually receives its sugar from the sources nearest to it. ∙Sugar from mesophyll cells or other sources must be loaded into sieve-tube members before it can be exported to sugar sinks. ∙In some plants, companion cells (transfer cells) have numerous ingrowths in their walls to increase the cell’s surface area and enhance the transfer of solutes between apoplast and symplast. ∙Downstream, at the sink end of the sieve tube, phloem unloads its sucrose.
Pressure flow ∙Pressure flow is the mechanism of translocation in angiosperms. ∙Phloem sap flows from source to sink at rates as great as 1 m/hr, faster than can be accounted for by either diffusion or cytoplasmic streaming. ∙Pressure flow in a sieve tube drives the bulk flow of phloem sap. ∙The closer the aphid’s stylet is to a sugar source, the faster the sap will flow and the greater its sugar concentration. ∙Genetic engineering of higher-yielding crop plants may depend on a better understanding of factors that limit bulk flow of sugars.
Chapter 37 Soil and Plant Nutrition
Soil texture ∙Texture of soil depends on the sizes of its particles. The size of the particles affects the availability of water, oxygen, and minerals. ∙Nutrition is the process by which an organism takes in and makes use of food substances. ∙Humus is the remains of dead organisms and other organic matter. ∙Soil Horizons aresoil layers. ∙Loams are the most fertile soil type, made up of roughly equal amounts of sand, silt, and clay.
Topsoil composition ∙A soil’s composition is made of inorganic and organic chemical components. ∙Topsoil’s ecosystem is made up with bacteria, fungus, protists, animals, and the roots of plants. ∙Cation exchange is a process in which positively charged minerals are made available to a plant when hydrogen ions in the soil displace mineral ions from the clay particles.
Conservation ∙The goal of soil conservation is to reduce damage. ∙Some agricultural practices can deplete the mineral content of soil, tax water reserves, and promote erosion. ∙Sustainable agriculture is a commitment embracing a variety of farming methods that are conservation minded, environmentally safe, and profitable. ∙No-till agriculture is a plowing technique that involves creating furrows, resulting in minimal disturbance of the soil. ∙Phytoremediation is an emerging nondestructive biotechnology that seeks to cheaply reclaim contaminated areas by taking advantage of some plant species’ ability to extract heavy metals and other pollutants from the soil and to concentrate them in easily harvested portions of the plant.
Macro and micronutrients ∙Macronutrients are elements that are required in large amounts: such as Carbon, Hydrogen, Oxygen, Nitrogen, and Phosphorus and Sulfur. ∙Micronutrients are elements required in small amounts, usually have catalytic functions as cofactors of enzymes. ∙Essential element is a chemical element required for the plant to grow from a seed and complete its life cycle, producing another generation in the form of seeds. ∙Hydroponic culture is a method in which plants are grown in minerals solutions rather than soil.
Table 37.1 Shows all the essential macronutrient and micronutrient elements in plants.
Healthy Phosphate-deficient Potassium-deficient Nitrogen-deficient Mineral deficiency ∙Deficiency of mobile nutrient usually affects older plants and for less mobile nutrient affects younger plants. ∙Deficiency of potassium, phosphorus, and nitrogen especially are the most common. Micronutrients are less common. ∙Moderation is important because overdose of many nutrients can be toxic to plants. Figure 37.7 The most common mineral deficiencies, with varying symptoms depending on species.
Genetic modification ∙Examples: resistance to aluminum toxicity, flood tolerance, and smart plants. ∙Rather than tailoring the soil to match the plant, genetic engineers are tailoring the plant to match the soil.
Atmosphere N2 N2 Atmosphere Soil Nitrate and nitrogenousorganiccompoundsexported inxylem toshoot system Nitrogen-fixingbacteria N2 Denitrifyingbacteria H+ (From soil) NH3 (ammonia) NH4+ Soil NO3– (nitrate) NH4+ (ammonium) Nitrifyingbacteria Ammonifyingbacteria Organicmaterial (humus) Root Figure 37.9 The roles of soil bacteria in the nitrogen nutrition of plants. Soil bacteria ∙Soil bacteria are decomposers. ∙Other soil bacteria, called rhizobacteria, derive their energy from the rhizosphere. Some rhizoshpere bacteria produce antibiotics. ∙Plants satisfy most of their huge needs for nitrogen from the bacterial decomposition of humus and the fixation of nitrogen.
Nitrogen fixing bacteria ∙Convert atmospheric N-2 to nitrogenous minerals that plants can absorb as a nitrogen source for organic synthesis. The most efficient mutualism between plants and nitrogen fixing bacterias occurs in the nodules formed by Rhizobium bacteria growing in the roots of legumes. These bacteria obtain sugar from the plant and supply the plant with fixed nitrogen. ∙A nodule is a swelling on the root of a legume. They are composed of plant cells that contain nitrogen-fixing bacteria of the genus Rhizobium. ∙A bacteriod is a form of the bacterium Rhizobium contained within the vesicles formed by the root cells of a root nodule. ∙Crop rotation is the practice of planting non-legumes one year and legumes in alternating years to restore concentrations of fixed nitrogen in the soil.
Fungi and plant nutrition ∙Mycorrhizae are mutualistic associations of fungi and roots. ∙There are two types of mycorrhizae: ectomycrrhizae and arbuscular mycorrhizae. ∙Ectomycorrhiaze is an association of a fungus with a plant root system in which the fungus surrounds the roots but does not cause invagination of the host cells’ plasma membrane. Arbuscular mycorrhizae is an association of a fungus with a plant root system in which the fungus causes the invagination of the host cells’ plasma membranes. ∙Both absorb water and minerals, which they supply to their plant hosts.
Epidermis Cortex (b) 10 m 2Endomycorrhizae. No mantle forms around the root, but microscopic fungal hyphae extend into the root. Within the root cortex, the fungus makes extensive contact with the plant through branching of hyphae that form arbuscules, providing an enormous surface area for nutrient swapping. The hyphae penetrate the cell walls, but not the plasma membranes, of cells within the cortex. Cortical cells Epidermis Cortex Mantle(fungalsheath) (a) 100 m aEctomycorrhizae. The mantle of the fungal mycelium ensheathes the root. Fungal hyphae extend from the mantle into the soil, absorbing water and minerals, especially phosphate. Hyphae also extend into the extracellular spaces of the root cortex, providing extensive surface area for nutrient exchange between the fungus and its host plant. Endodermis Fungalhyphae Vesicle Endodermis Casparianstrip Roothair Fungalhyphaebetweencorticalcells Arbuscules (LM, stained specimen) Mantle(fungal sheath) (colorized SEM) Figure 37.12 Two types of mycorrhizae.
Unusual adaptations Epiphytes grow on the surface of other plants but acquire water and minerals from rain. Parasitic plants absorb nutrients from host plants. Carnivorous plants supplement their mineral nutrition by digesting animals.
Chapter 38 Angiosperm Reproduction and Biotechnology
Life cycles ∙The life cycles of plants are characterized by an alternation of generations, in which multi-cellular haploid (n) and diploid (2n) generations take turns producing each other. ∙The diploid plant, which is called the sporophyte, produces haploid spores by meiosis. These spores divide by mitosis creating multi-cellular gametophytes. Once these are fertilized they become diploid.
Structure and function ∙The flower is the reproductive part of a plant. ∙There are five main parts to a flower: sepal, petal, receptacle, carpel, and stamen. ∙Sepals and petals are both sterile parts of the flower. They can be used to collect water or attract pollinators. ∙The receptacle is where the flower is attached. It supports the flower and provides it with the nutrients it needs. ∙The carpel, also called the pistil, consist of the stigma, the style, and the ovary. The stigma is a sticky structure that collects the pollen. The style is the long neck that connects the stigma to the ovary. The ovary contains the ovules. This is where fertilization occurs. ∙The stamen includes the anther and the filament. The anther is where the micro- sporangia are located. The microsporangium are the pollen sacs. This is where the pollen is created.
Stigma Carpel Anther Stamen Style Filament Ovary Sepal Petal Receptacle Figure 38.2 An idealized flower.
Male gametophytes ∙Each anther contains four pollen sacs, also known as microsporangia. These contain many diploid cells. ∙Each cell undergoes meiosis forming four haploid microspores. ∙These will eventually turn to haploid male cells. Each then goes into mitosis to produce a mal gametophyte which consist of only two cells. ∙After the male gametophyte is formed it matures and travels up the spore wall. ∙This cell will release the pollen into the pollen tube to be delivered to the female gametophyte.
Female gametophytes ∙There are fifteen variations on how the female gametophyte is created. ∙One way is one that takes place only in the carpel’s ovary. ∙Each ovule consist of one megasporocyte which will expand and undergo meiosis. Only one will survive. ∙The surviving one will continue to grow while its nucleus undergoes mitosis three times without cytokinesis. ∙This will result in one large cell with eight haploid nuclei. This cell will now be taken to the embryo sac to be fertilized. Two cells called synergids flank the egg cell and help guide pollen to the pollen tube.
Figure 38.3 The development of male and female gametophytes in angiosperms. Pollen sac (microsporangium) (b) Development of a female gametophyte (embryo sac) Mega- sporangium (a) Development of a male gametophyte (pollen grain) Within the ovule’s megasporangium is a large diploid cell called the megasporocyte (megaspore mother cell). 1 Ovule Mega- sporocyte Micro- sporocyte Each one of the microsporangia contains diploid microsporocytes (microspore mother cells). MEIOSIS 1 MEIOSIS Integuments Micropyle Surviving megaspore Micro- spores (4) Female gametophyte (embryo sac) Each microsporo- cyte divides by meiosis to produce four haploid microspores, each of which develops into a pollen grain. MITOSIS 2 Antipodel Cells (3) Each of 4 microspores Ovule MITOSIS Polar Nuclei (2) The megasporocyte divides by meiosis and gives rise to four haploid cells, but in most species only one of these survives as the megaspore. 2 Generative cell (will form 2 sperm) Egg (1) Male Gametophyte (pollen grain) Integuments Synergids (2) Key to labels Embryo sac Nucleus of tube cell KEYto labels Haploid (2n) 100 m 20 m Diploid (2n) Three mitotic divisions of the megaspore form the embryo sac, a multicellular female gametophyte. The ovule now consists of the embryo sac along with the surrounding integuments (protective tissue). 3 Haploid (2n) Ragweedpollengrain A pollen grain becomes a mature male gametophyte when its generative nucleus divides and forms two sperm. This usually occurs after a pollen grain lands on the stigma of a carpel and the pollen tube begins to grow. (See Figure 38.2b.) 3 Diploid (2n) 75 m
Pollination ∙Pollination is defined by the transfer of pollen from an anther to a stigma. ∙Wind pollinated species like grasses and most trees create more pollen to compensate for the random dispersion of the pollen to the plants. This is why the air is full of pollen during the spring. ∙Another form of pollination is through the water for plants that grow there. ∙Most angiosperms use insects to pollinate them. This method uses an insect to directly carry pollen from one flower to another. ∙If a plant is fertilized twice, the extra pollen connects to extra nuclei to form endosperm. This works as a food storing tissue of the seed. This is called double fertilization.
Seeds ∙After double fertilization each ovule develops into a seed while each ovary develops into the fruit holding the seed or seeds. The fruit then can act as a sugar storage. ∙Once the seed is matured it dehydrates itself and enters dormancy. ∙When a seed is in dormancy, its metabolic processes are basically non-existent. ∙The seed is then enclosed by a hard coating. The seed consist of the radicle, the seed coat, the epicotyl, the hypocotyl, and the cotyledons. ∙Seeds can stay dormant for a long time. They won’t germinate until the conditions are right or some environmental change causes it to leave dormancy. ∙Seeds will germinate differently depending on their location. Seeds in forests where fires are common will not germinate until after a fire has cleared away all the competing vegetation.
Seed coat Epicotyl Hypocotyl Radicle Cotyledons Figure 38.8 Common garden bean, a eudicot with thick cotyledons. The fleshy cotyledons store food absorbed from the endosperm before the seed germinates. Germination ∙A seed will germinate after it starts to take in water. The seed will expand from the water than bust the outer coat letting the plant begin growth. ∙The first organ to emerge is the radicle which is the root. ∙Next the hypocotyl will emerge out of the soil and reacting to light straighten. ∙Lastly leaves will grow on the end of the hypocotyl.