Download
1 / 1
10 likes | 139 Views
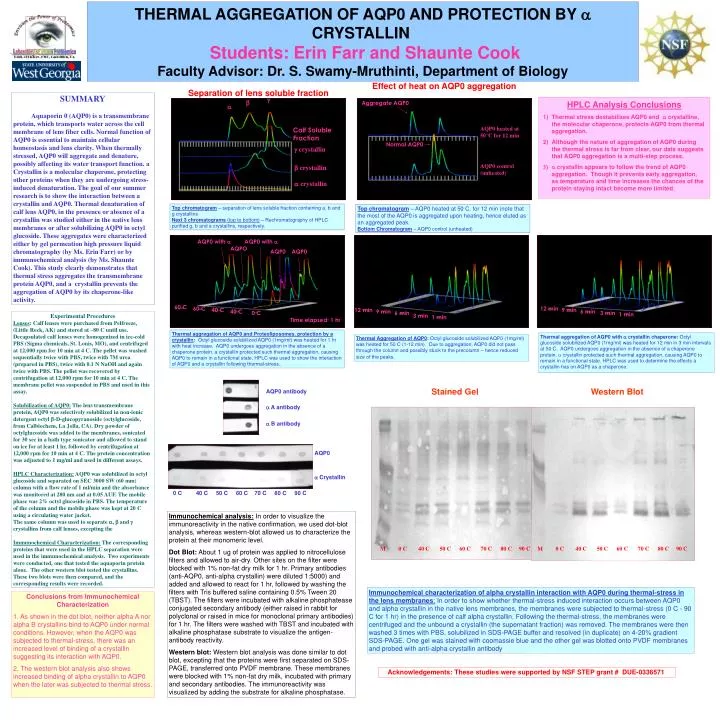
THERMAL AGGREGATION OF AQP0 AND PROTECTION BY a CRYSTALLIN Students: Erin Farr and Shaunte Cook Faculty Advisor: Dr. S. Swamy-Mruthinti, Department of Biology. Effect of heat on AQP0 aggregation. Separation of lens soluble fraction. SUMMARY

E N D
THERMAL AGGREGATION OF AQP0 AND PROTECTION BY a CRYSTALLINStudents: Erin Farr and Shaunte CookFaculty Advisor: Dr. S. Swamy-Mruthinti, Department of Biology Effect of heat on AQP0 aggregation Separation of lens soluble fraction SUMMARY Aquaporin 0 (AQP0) is a transmembrane protein, which transports water across the cell membrane of lens fiber cells. Normal function of AQP0 is essential to maintain cellular homeostasis and lens clarity. When thermally stressed, AQP0 will aggregate and denature, possibly affecting its water transport function. a Crystallin is a molecular chaperone, protecting other proteins when they are undergoing stress-induced denaturation. The goal of our summer research is to show the interaction between a crystallin and AQP0. Thermal denaturation of calf lens AQP0, in the presence or absence of a crystallin was studied either in the native lens membranes or after solubilizing AQP0 in octyl glucoside. These aggregates were characterized either by gel permeation high pressure liquid chromatography (by Ms. Erin Farr) or by immunochemical analysis (by Ms. Shaunte Cook). This study clearly demonstrates that thermal stress aggregates the transmembrane protein AQP0, and a crystallin prevents the aggregation of AQP0 by its chaperone-like activity. g b Aggregate AQP0 • HPLC Analysis Conclusions • Thermal stress destabilizes AQP0 and a crystalline, the molecular chaperone, protects AQP0 from thermal aggregation. • Although the nature of aggregation of AQP0 during the thermal stress is far from clear, our data suggests that AQP0 aggregation is a multi-step process. • a crystallin appears to follow the trend of AQP0 aggregation. Though it prevents early aggregation, as temperature and time increases the chances of the protein staying intact become more limited. a AQP0 heated at 50°C for 12 min Calf Soluble Fraction Normal AQP0 g crystallin AQP0 control (unheated) b crystallin a crystallin Top chromatogram – separation of lens soluble fraction containing a, b and g crystallins Next 3 chromatograms (top to bottom) – Rechromatography of HPLC purified g, b and a crystallins, respectively. Top chromatogram – AQP0 heated at 50 C. for 12 min (note that the most of the AQP0 is aggregated upon heating, hence eluted as an aggregated peak. Bottom Chromatogram –AQP0 control (unheated) AQP0 with a AQP0 with a AQPO AQP0 AQP0 60 C 60 C 12 min 40 C 12 min 9 min 40 C 9 min 6 min 0 C 6 min 3 min 1 min Experimental Procedures Lenses: Calf lenses were purchased from Pelfreeze, (Little Rock, AK) and stored at –80 C until use. Decapsulated calf lenses were homogenized in ice-cold PBS (Sigma chemicals, St. Louis, MO), and centrifuged at 12,000 rpm for 10 min at 4 C. The pellet was washed sequentially twice with PBS, twice with 7M urea (prepared in PBS), twice with 0.1 N NaOH and again twice with PBS. The pellet was recovered by centrifugation at 12,000 rpm for 10 min at 4 C. The membrane pellet was suspended in PBS and used in this assay. Solubilization of AQP0: The lens transmembrane protein, AQP0 was selectively solubilized in non-ionic detergent octyl b-D-glucopyranoside (octylglucoside, from Calbiochem, La Jolla, CA). Dry powder of octylglucoside was added to the membranes, sonicated for 30 sec in a bath type sonicator and allowed to stand on ice for at least 1 hr, followed by centrifugation at 12,000 rpm for 10 min at 4 C. The protein concentration was adjusted to 1 mg/ml and used in different assays. HPLC Characterization: AQP0 was solubilized in octyl glucoside and separated on SEC 3000 SW (60 mm) column with a flow rate of 1 ml/min and the absorbance was monitored at 280 nm and at 0.05 AUF. The mobile phase was 2% octyl glucoside in PBS. The temperature of the column and the mobile phase was kept at 20 C using a circulating water jacket. The same column was used to separate a, b and g crystallins from calf lenses, excepting the Immunochemical Characterization: The corresponding proteins that were used in the HPLC separation were used in the immunochemical analysis. Two experiments were conducted, one that tested the aquaporin protein alone. The other western blot tested the crystallins. These two blots were then compared, and the corresponding results were recorded. 3 min 1 min Time elapsed: 1 hr Thermal aggregation of AQP0 and Proteoliposomes, protection by a crystallin: Octyl glucoside solubilized AQP0 (1mg/ml) was heated for 1 hr with heat increase. AQP0 undergoes aggregation in the absence of a chaperone protein. a crystallin protected such thermal aggregation, causing AQP0 to remain in a functional state. HPLC was used to show the interaction of AQP0 and a crystallin following thermal-stress.. Thermal aggregation of AQP0 with a crystallinchaperone: Octyl glucoside solubilized AQP0 (1mg/ml) was heated for 12 min in 3 min intervals at 50 C. AQP0 undergoes aggregation in the absence of a chaperone protein. a crystallin protected such thermal aggregation, causing AQP0 to remain in a functional state. HPLC was used to determine the effects a crystallin has on AQP0 as a chaperone. Thermal Aggregation of AQP0: Octyl glucoside solubilized AQP0 (1mg/ml) was heated for 50 C (1-12 min). Due to aggregation, AQP0 did not pass through the column and possibly stuck to the precolumn – hence reduced size of the peaks. Stained Gel Western Blot AQP0 antibody a A antibody a B antibody AQP0 a Crystallin 0 C 40 C 50 C 60 C 70 C 80 C 90 C Immunochemical analysis: In order to visualize the immunoreactivity in the native confirmation, we used dot-blot analysis, whereas western-blot allowed us to characterize the protein at their monomeric level. Dot Blot: About 1 ug of protein was applied to nitrocellulose filters and allowed to air-dry. Other sites on the filter were blocked with 1% non-fat dry milk for 1 hr. Primary antibodies (anti-AQP0, anti-alpha crystallin) were diluted 1:5000) and added and allowed to react for 1 hr, followed by washing the filters with Tris buffered saline containing 0.5% Tween 20 (TBST). The filters were incubated with alkaline phosphatease conjugated secondary antibody (either raised in rabbit for polyclonal or raised in mice for monoclonal primary antibodies) for 1 hr. The filters were washed with TBST and incubated with alkaline phosphatase substrate to visualize the antigen-antibody reactivity. Western blot: Western blot analysis was done similar to dot blot, excepting that the proteins were first separated on SDS-PAGE, transferred onto PVDF membrane. These membranes were blocked with 1% non-fat dry milk, incubated with primary and secondary antibodies. The immunoreactivity was visualized by adding the substrate for alkaline phosphatase. M 0 C 40 C 50 C 60 C 70 C 80 C 90 C M 0 C 40 C 50 C 60 C 70 C 80 C 90 C Immunochemical characterization of alpha crystallin interaction with AQP0 during thermal-stress in the lens membranes: In order to show whether thermal-stress induced interaction occurs between AQP0 and alpha crystallin in the native lens membranes, the membranes were subjected to thermal-stress (0 C - 90 C for 1 hr) in the presence of calf alpha crystallin. Following the thermal-stress, the membranes were centrifuged and the unbound a crystallin (the supernatant fraction) was removed. The membranes were then washed 3 times with PBS, solubilized in SDS-PAGE buffer and resolved (in duplicate) on 4-20% gradient SDS-PAGE. One gel was stained with coomassie blue and the other gel was blotted onto PVDF membranes and probed with anti-alpha crystallin antibody Conclusions from Immunochemical Characterization 1. As shown in the dot blot, neither alpha A nor alpha B crystallins bind to AQP0 under normal conditions. However, when the AQP0 was subjected to thermal-stress, there was an increased level of binding of a crystallin suggesting its interaction with AQP0. 2. The western blot analysis also shows increased binding of alpha crystallin to AQP0 when the later was subjected to thermal stress. Acknowledgements: These studies were supported by NSF STEP grant # DUE-0336571






















