Download
1 / 10
100 likes | 243 Views
Large Punctuational Contribution of Speciation to Evolutionary Divergence at the Molecular Level. Jaksottaisen tasapainon evoluutiomalli. Jatkuvan, tasaisen morfologisen muutoksen kilpailija, ei gradualismin (hidas, vähittäinen muutos): kladogeneesi vs. anageneesi

E N D
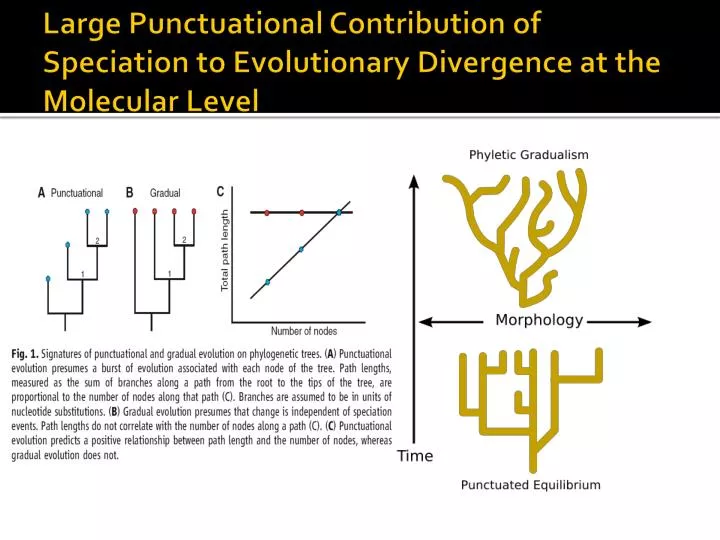
Large Punctuational Contribution of Speciation to Evolutionary Divergence at the Molecular Level
Jaksottaisen tasapainon evoluutiomalli • Jatkuvan, tasaisen morfologisen muutoksen kilpailija, ei gradualismin (hidas, vähittäinen muutos): kladogeneesi vs. anageneesi • Jaksottaisia kehityspyrähdyksiä ja pitkiä suvantovaiheita, jolloin populaatiot tasapainossa ympäristön valintapaineiden kanssa • Pyrähdyksissä populaatio siirtyy tasapainosta toiseen • Muutokset lajitasolla nopeita, fossiiliaineistoon jää vähän jälkiä muutosten vaiheista välimuotofossiilien löytyminen epätodennäköistä
PE vastaa n. 20% evoluution evoluutiosta kun käytetyssä puussa on yli 35 osaa
Ratkaistu betan arvo oli keskimäärin 0.0078 + 0.0017, absoluuttinenarvoriippuugeenienevoluutionopeudesta, joitaanalysoiduissapuissakäytettiin • Kun beta = 0, rhon arvo on 0 (odotettu korrelaatio polkujen pituuden ja haarautumispisteiden välillä) • Sigma n on puun solmukohtien määrän varianssi • Sigma g on graduaalisten vaikutusten varianssi • Sigma g,n on g:n (g komponentti) ja n:n (kuljetun polun solmukohtien lkm =0) välinen kovarianssi • Alunperin 35 puutasisälsi b>0: Average correlation between path lengths and nodes ranges from 0.22 to 0.69 with a mean of r = 0.42 + 0.019; punctuational effects contributed between 4.8 and 48% (mean r^2 = 0.42^2 = 0.18) of the deviation from molecular clock–like behavior in these trees.
Erratum • ”…previously undescribed bias that affects Bayesian posterior distributions of phylogenetic trees derived from Markov chain Monte Carlo methods. The bias arises when species are closely related and thus the phylogenetic signal is difficult to detect. The bias manifests itself as a tendency in the posterior sample toward asymmetrically branching trees with short but variable branch lengths. Under these circumstances, the posterior distribution of trees can support the inference of punctuational evolution even when no such effect is present.” • 11/120 käytetystäfylogeneettisestädatastakärsiivirheestä, eisiltimuutapäätelmiä: 27 ± 4.7% (aiemmin 35 ± 4.8%) puista sisältää jaksottaisen evoluution aiheuttamia muutoksia
Tulkinta • Jos fylogeneettisen puun koko olisi ääretön, arviolta 16 ± 5.4% kaikista geneettisistä muutoksista tapahtuisi jaksottaisen tasapainon mallin mukaisesti • Kasveilla yleisyys 43.5 ± 10.0% • Sienillä 60.0 ± 22% • Eläimillä 18.0 ± 4.9%, alle ½x pienempi tdn kuin em. • ”The size of the punctuational effect predicts departures from a molecular clock-like mode of evolution”, ”By masquerading as gradual changes, punctuational effects will bias estimates of dates derived from molecular clocks that assume gradual evolution, making them occur too far in the past.”
Vaihtoehtoja • Rather than measuring a burst of evolution, we are detecting a generalized increase in the rate of evolutionafter speciation • A second general mechanism for divergence is adaptive evolution as species invade new niches • Sympatric speciation is far more common than previously believed: arises from specific and often rapid isolating mechanisms operating in small groups • Isolaatiomekanismit: esim. käytöstä muuttavat mutaatiot, kromosomien uudelleen järjestäytymiset, ploidian muutokset ja hybridisaatio usein tuottavat populaation sisäisiä pieniä ryhmiä jotka eivät lisäänny alkuperäisen muodon kanssa
Lajiutuminen adaptiivisen radiaation kautta • 1. Species A migrates from the mainland to the first island. • 2. Isolated from the mainland, species A evolves to species B. • 3. Species B migrates to the second island. • 4. Species B evolves in species C. • 5. Species C recolonizes the first islands, inable to reproduce with species B. • 6. Species C migrates to the third island. • 7. Species C evolves into species D. • 8. Species D migrates to the first and second island. • 9. Species D evolves to species E. • The story could go on and on, until a large diversity is reached.