Download

1 / 17
170 likes | 354 Views
El splicing de oskar mRNA esta acoplado con su localización citoplasmática. Y14 y Mago se colocalizan con oskar m RNA en el polo posterior del oocito de D. melanogaster .

E N D
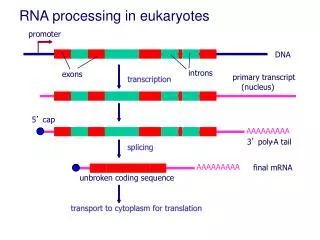
El splicing de oskar mRNA esta acoplado con su localización citoplasmática
Y14 y Mago se colocalizan con oskarmRNA en el polo posterior del oocito de D. melanogaster. • La correcta localización del transcrito de oskar es esencial para el desarrollo del abdomen (y para la línea germinal).
Splicing del primer intrón de oskar mRNA es esencial para su correcta localización. ¿Contradicción con la suficiencia de la región 3’ UTR para la correcta localización? Puede que los anteriores trabajos estuviesen afectados por transcritos de oskar endógenos.
Efectos de diferentes oskar mRNA con intrones delecionados (en oskA87/Df(3R)pXT103)
Efectos de diferentes oskar mRNA con intrones delecionados (en oskA87/Df(3R)pXT103) • oskΔi(1,2,3)transporte correcto, pero difuso a partir del estadío 8 aunque hay la misma cantidad de transcrito. 2/3 de los huevos no eclosionan. Poca traducción. • Transcritos transgénicos con i1 [oskWT, oskΔi2 , oskΔi3 y oskΔi(2,3)] correctamente localizados. Suficiente para que Y14 se asocie con el transcrito.
Transcritos transgénicos sin i1 [oskΔi1 oskΔi(1,2) oskΔi(1,3) ] no se localiza en el polo posterior en los estadios 9 y 10 aunque el splicing de i2 y i3 sea correcto.
Conclusión i1 tiene una función diferente de i2 y i3 en la localización del transcrito. Hay 2 posibilidades: • i1 contiene información específica de secuencia. • El splicing en la posición i1 es esencial para la localización de oskar mRNA.
Utilizando el transgénico osk(i3 in i1) se vio que se localizaba correctamente con Y14. Así pues el reclutamiento de Y14 es independiente de la secuencia del intrón.
Revisión de los trabajos con RNA híbrido lacZ-osk3’UTR Puede que oskar RNA endógeno afecte a los resultados. La localización de lacZ-osk3’UTR en oskA87/Df(3R)pXT103 no es posible porque la oogenesis se detiene en sus primeros estadios. Se utilizaron: • oskWT localizacion correcta • oskΔi(1,2,3) deslocalizado • osk84 localización correcta
Conclusiones • LacZ-osk3’UTR necesita una fuente endógena de oskar mRNA para su localización en el polo posterior del oocito. • La localización es independiente de la proteína Oskar. • La localización de lacZ-osk3’UTR en el polo posterior se debe probablemente a que se une a los complejos mRNP. • La región 3’UTR es esencial, ya que los trancritos`sin 3’UTR no se localizan.
Solo la deposición del EJC en la primera unión exón-exón es esencial, aunque EJC se une a todas. Se requiere una señal de EJC y una posición concreta. Papel estructural del heterodímero Y14-Mago nashi, y de eIF4AIII y Barentsz.
Conclusiones • Que el splicing, y Y14 y Mago nashi sean esenciales para su localización, demuestra que el splicing y la localización están acopladas por la deposición dependiente del splicing del EJC.
Propuesta de función del EJC • Especificar la estructura del mRNP según sea su localización citoplásmica. • En humanos la señalización con EJC permite reconocer codones de parada prematuros provocando la activación de NMD (degradación de mRNA). • En Drosophila factores de NMD y EJC están conservados, pero la terminación prematura no depende de EJC ni de la posición del intrón. Sugiere un complejo conservado con diferentes funciones.
No quiere decir que el EJC no este implicado en la localización citoplásmica de mRNA en vertebrados, ya que se ha detectado Barentsz en neuronas del hipocampo. • Se podrá utilizar para determinar si el transporte de otros mRNAs es mediante EJC y para determinar la importancia de la conservación del EJC.