Download
1 / 49
490 likes | 661 Views
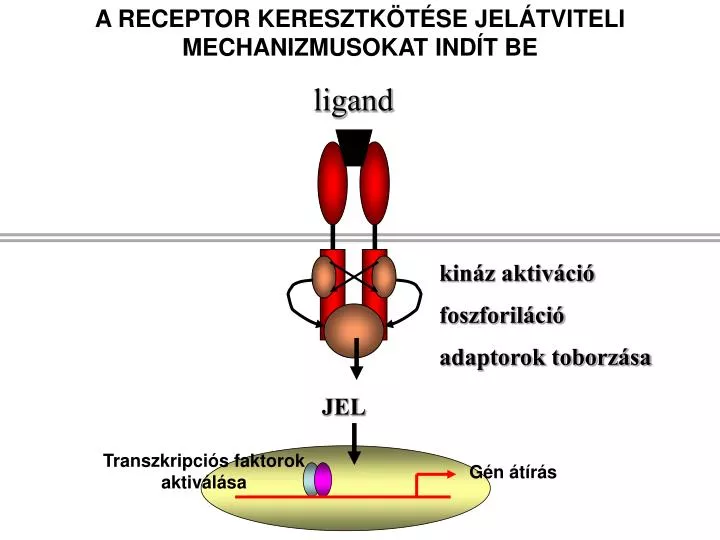
A RECEPTOR KERESZTKÖTÉSE JELÁTVITELI MECHANIZMUSOKAT INDÍT BE. ligand. k ináz aktiváció. foszforiláció. adaptorok toborzása. Transzkripciós faktorok aktiválása. Gén átírás. JEL. a. a. antig e n binding. V. V. V. V. L. L. mIg molecule. Kin ases. H. H. Signal transduction. b.

E N D
A RECEPTOR KERESZTKÖTÉSE JELÁTVITELI MECHANIZMUSOKAT INDÍT BE ligand kináz aktiváció foszforiláció adaptorok toborzása Transzkripciós faktorok aktiválása Gén átírás JEL
a a antigen binding V V V V L L mIg molecule Kinases H H Signal transduction b SHP-1 a Syk Phosphatases Btk PLC Vav Lyn HS1 Adaptors + substrates SLP-65/BLNK THE IgM B-CELL RECEPTOR Ig-a/Ig-b heterodimer
Ig domén + CHO a b ITAM ITAM Y Y Y Y A B-SEJT RECEPTOR FELÉPÍTÉSE Ig-a/CD79a Ig-b/CD79b ITAM: YxxLx7YxxI ITAM: Immunoreceptor Tyrosine-based Activation Motif
Ag 2. Src-family kináz activáció és ITAM foszforiláció 3. Syk toborzás és aktiváció 4. SLP foszforiláció +Ca felszabadulás Syk Syk Lyn Lyn SLP P P P P P P P Kalcium felszabadulás = ITAM B-SEJT RECEPTOR SZIGNÁLÚTVONAL 1. kereztkötés
A LIMFOCITA AKTIVÁCIÓ KINETIKÁJA Nyugvó limfocita G0 Nyugvó limfocita G0 Ko-receptor Adhéziós molekulák Citokinek 2. jel Effektor sejtMemóris sejt Transzport Membrán változások RNS és fehérje szintézis proliferáció DNS szintézis Limfoblaszt PTK aktiváció RNS szintézis szabad Ca++ Fehérje szintézis fehérje foszforiláció DNS szintézis Nyugvó limfocita G0 0 10sec 1min 5min 1hr 6 hrs 12 hrs 24 hrs ANTIGEN SIGNAL1.
AZ ELLENANYAGOK SZERKEZETE Nehéz lánc (H) VH VL CH Könnyű lánc (L) CL KOMPLEMENT AKTIVÁCIÓ SEJTHEZ KÖTŐDÉS LEBOMLÁS TRANSZPORT
AZ ELLENANYAG SOKFÉLESÉG GENETIKAI HÁTTERE VH VH VL VL S – S S – S A sokféleség kialakulásának mechanizmusa? Más szabályok a variábilis és konstans régiók kialakulására? Szimmetrikus molekula két azonos VH és VL mindkét kromoszóma ugyanazt a szekvenciát kódolja?
AZ IMMUNOGLOBULINOK JELLEGZETES AMINOSAV SZERKEZETE Mieloma multiplex Plazmasejt tumorok – tumorsejtek a csontvelőben Monoklonális eredetű emberi immunoglobulinok a szérumban(50-100mg/ml) Rodney Porter és Gerald Edelman 1959 – 1960 fehérje tisztítás L H Redukció 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 Variábilis Constans Gél elektroforézis 50 kDa Nehéz lánc 25 kDa Könnyű lánc
A könnyű lánc egyes részei variábilisak, mások konzerváltabbak: Hipervariábilis és framework régiók
A MOLEKULÁRIS BIOLÓGIAI DOGMÁJA AZ IMMUNOGLOBULIN SZEKVENCIA JELLEGZETESSÉGE 1 GÉN = 1FEHÉRJE ELMÉLETEK 1 GÉN magas szomatikus mutációs ráta a V régióban Gén V C Sok GÉN (10000 – 100000) Protein V1 C V2 C V3 C
AZ IMMUNOGLOBULINOK MOLEKULÁRIS GENETIKÁJA Hogyan magyarázható az ellenanyagok kettős funkciója? Dreyer & Bennett feltételezése (1965) Egy adott izotípusú ellenanyag valószínűleg: • Az egyetlen C régiót kódoló gén a csíravonalban el van választva a V régió génektől • A V génekből többféle áll rendelkezésre • Feltételezhető egy mechanizmus, amely a V és C géneket fúzionálja egy teljes immunoglobulin génné A feltételezés ellentétben állt az akkor elfogadott nézettel, amennyiben a DNS (genetikai állomány) egy adott egyed minden sejtjében azonos
A csíravonalban egyetlen Cgén van (L-lánc), amely a sok V géntőltávol helyezkedik V V V V V V V V V C V V V C A B-sejtekben a V és C géneket valamilyen mechanizmus egy teljes immunoglobulin génné fúzionálja V V A Dreyer - Bennett hipotézis igazolása Módszer a sok V gén létezésének és a V – C gén átrendeződés igazolására
V V Csíravonal DNS V V V V V V V C V V V C Átrendezett DNS V V Módszer • Megközelítés: • Specifikus cDNS próbák a különböző V régiók és a C régiók elkülönítéséhez • DNS restrikciós enzimek a DNS fragmentáláshoz • A csíravonalnak (pl. méhlepény)és az érett B-sejteknek (pl. plazmocitóma/mielóma) a DNS-ei eltérőek
Kb V-próba 6,0 V C-próba 4,0 C 1,5 VC 6.0 Kb 4.0 Kb V V C C 1.5. Kb Susumi Tonegawa kísérlete 1975 DNS-kivonás Restrikciós enzim hasítás Gél elektroforézis Hibridizáció B-sejt Méhlepény B-sejt
The actual Tonegawa experiment 1. DNS BamHI emésztés 2. Elektroforézis 3. Gélszeletek készítése (MW Szerint) 4. DNS extrakció 5. Hibridizáció 125I-jelölt Kappa lánc RNS-sel 6. RNAse kezelés. (DNS-RNS hibrid RNAse rezisztens 7. Precipitáció (DNS-RNS hibrid igen a nem kötődött próba nem) 8. Radioaktivitás mérése
KÖVETKEZTETÉS Gén A V és C gének csak a B-sejtekben kerülnek egymás mellé V V V V V V V C C B-SEJT GÉN SZEGMENSEK SZOMATIKUS ÁTRENDEZŐDÉSE EGY GÉNNÉ Fehérje Sok variábilis gén van, de csak egy konstans gén CSÍRAVONAL
L VL JL CL L VL CL ~ 95as ~ 100as ~ 95as ~ 100as VL CL L Az aminosavak egy része a kis számú J (Joining) régiókból származik ~ 208as Az Ig gének szekvenálása tovább bonyolította a képet A csíravonal VL gének szerkezete hasonló volt a Vkés and Vlkönnyű láncok esetében A csíravonal és az átrendezett DNS nem volt azonos Honnan származik a 13 extra aminosav?
L VL JL CL Az Ig H lánc további sokféleséggel jellemezhető DH JH L VH CH A nehéz lánc a JH és CH gének közötti szakaszon további (0 – 8) aminosavat tartalmaz Ezek a D (DIVERSITY) régióból származnak A nehéz lánc kialakulásához 2 rekombinációs folyamatra van szükség JH to DH, VH to JHDH A könnyű lánc kialakulásához 1 rekombinációs folyamatra van szükség : VL to JL
Gén szegmens Könnyű lánc Nehéz lánc kappa lambda Variábilis (V) 40 30 65 Diverzitás (D) 0 0 27 Kapcsoló (J) 5 4 6 AZ IMMUNOGLOBULIN POLIPEPTID LÁNCOKAT TÖBB GÉN SZEGMENS KÓDOLJA AZ IMMUNOGLOBULIN GÉN SZEGMENSEK ELRENDEZŐDÉSE 2 kromoszóma kappa könnyű lánc gén szegmensek 22 kromoszóma lambda könnyű lánc gén szegmensek 14 kromoszóma nehéz lánc gén szegmensek AZ IMMUNOGLOBULIN GÉN SZEGMENSEK SZÁMA
A KAPPA (κ) LÁNC GÉN SZEGMENSEK SZOMATIKUS ÁTRENDEZŐDÉSE B-sejt2 5 Jκ 40 Vκ Csíravonal Vκ Jκ Jκ Jκ Jκ Vκ Vκ Vκ Vκ Vκ Vκ Vκ B-limfocita fejlődés során Jk Jκ Jκ Jκ B-sejt1 Jκ DNS
pA Vκ P Vκ Vκ Vκ Vκ J J J J J J E E VκJκ Primer RNSátirat Cκ Cκ Cκ Cκ mRNA AAAA Transzláció Fehérje A K-LÁNC KIFEJEZŐDÉSE
A B sejt fejlődés során VH VH VH D D D D D D D D JH JH VH JH JH VH VH A VH GÉN SZEGMENSEK SZOMATIKUS ÁTRENDEZŐDÉSE 65 VH 27 D 6 JH VH VH JH JH JH JH
VH JH D V-Domének C-Domének VL JL VH-D-JH VL-JL AZ ANTIGÉN RECEPTOROK SOKFÉLESÉGE
AZ IMMUNOGLOBULIN GÉN SZEGMENSEK ÁTRENDEZŐDÉSÉNEK SORRENDJE D – J rekombináció Pót könnyű lánc V – DJ rekombináció V – J rekombináció VDJ –δ transzkripció δ transzláció VJ – (vagy VJ - ) transzkripció vagy transzláció B-sejt mIgD mIgM
A kombinációs sokféleség becslése A funkcionális V, D és J gének száma: 65 VH x 27 DH x 6JH = 10, 530 kombináció 40 Vk x 5 Jk = 200kombináció 30 Vl x 4 Jl = 120 kombináció = 320különböző könnyű lánc Amennyiben a H és L láncok véletlenszerűen párosodnak mint H2L2 19,440 x 265 = 3,369,600lehetőség Csak a KOMBINÁCIÓSsokféleség A valóságban bizonyos H + L kombinációk nem fordulnak elő, mert instabilak Bizonyos V és J gének gyakrabban fejeződnek ki, mint mások A KAPCSOLÁSI sokféleség tovább növeli a szekvenciák számát A POTENCIÁLIS B-SEJT KÉSZLET KIALAKULÁSA
A SZOMATIKUS GÉN ÁTRENDEZŐDÉS EREDMÉNYE • A gén szegmensek kombinációjanagy számú, eltérő variábilis régióval rendelkező nehéz (H) és könnyű (L) láncot eredményez, amelyek egy adott egyedben a különböző B-sejt klónokban fejeződnek ki • Az egyik kromoszómán létrejött sikeres gén átrendeződés gátolja a gén átrendeződést a másik kromoszómán • ALLÉL KIZÁRÁS • 3. Egy B-sejt csak egy típusú nehéz (H) és egy típusú könnyű (L) láncot termel • ELKÖTELEZŐDIK EGY TÍPUSÚ ANTIGÉN KÖTŐHELY LÉTREHOZÁSÁRA • 4. A teljes B sejt készlet különbözőképpen átrendezett immunoglobulin gének termékeit kifejező egyedi B-sejtekből áll
A B-SEJT ANTIGÉN FELISMERŐ RECEPTOR ÉS AZ ELLENANYAGOK SOKFÉLESÉGÉT MEGHATÁROZÓ GENETIKAI MECHANIZMUSOK • Szomatikus génátrendeződés • A variábilis domének kialakulása a VJ (L-lánc) és a VDJ (H-lánc) gének rekombinációjával • Hogyan képes a B sejt csak egyfajta könnyű és egyfajta nehéz láncot kifejezni? • Annak ellenére, hogy minden B sejtben egy apai és egy anyai Ig lókusz is jelen van • A gének nagy része ko-dominánsan kifejeződik • Hogyan történik a B sejtekben az egyik allél aktiválása és a másik gátlása? ALLÉL KIZÁRÁS • A sikeres génátrendeződés gátolja a rekombinációt a másik kromoszómán • Csak egyféle H- és L-lánc szintetizálódik • A sejtfelszínre 2 H- és 2 L-láncból álló dimerek kerülnek ki AZ ANTIGÉN JELENLÉTÉTŐL FÜGGETLEN FOLYAMATOK A B-SEJT ÉRÉS SORÁN A CSONTVELŐBEN MENNEK VÉGBE
How does somatic gene rearrangement (recombination) work? • How is an infinite diversity of specificity generated from finite amounts of DNA? • Combinatorial diversity • 2. How do V region find J regions and why don’t they join to C regions?12-23 rule -Special - Recobnitation Signal Sequences (RSS) - Recognized by Recombination Activation Gene coded proteins (RAGs) PALINDROMIC SEQUENCES HEPTAMER CACAGTG CACAGTG GTGACAC GTGACAC NONAMER ACAAAAACC GGTTTTTGT TGTTTTTGG CCAAAAACA
Somatic recombination to generate antibody diversity
Súlyos kombinált immundrficiencia Severe combined immunodeficiency syndrome (SCID). Omen syndrome RAG deficiency Korai manifesztáció Erős hasmenés Vörös foltok a testen Opportunista fertőzések (Candida albicans, Pneumocystis carnii pneumonia) Nyirokcsompk nem tapinthatók
Vl Jl 7 23 12 7 9 9 Vk JH Jk 9 9 7 12 23 23 7 7 9 D 12 7 7 12 9 9 VH 9 7 23 V, D, J flanking sequences Sequencing upstream and downstream of V, D and J elements revealed conserved sequences of 7, 23, 9 and 12 nucleotides in an arrangement that depended upon the locus
HEPTAMER - Always contiguous with coding sequence NONAMER - Separated fromthe heptamer by a 12 or 23 nucleotide spacer JH JH 9 9 23 23 7 7 D D 12 12 7 7 7 7 12 12 9 9 9 9 VH VH 9 9 7 7 23 23 Recombination signal sequences (RSS) 12-23 RULE – A gene segment flanked by a 23mer RSS can only be linked to a segment flanked by a 12mer RSS
12-mer = one turn 23-mer = two turns Intervening DNA of any length 23 12 V 7 9 7 D J 9 A 12-23 szabály molekuláris háttere
V4 V5 V3 V1 V3 V4 V2 V6 V2 V5 V6 V7 V8 V7 9 V9 D J V8 V9 9 23-mer • Heptamers and nonamers align back-to-back • The shape generated by the RSS’s acts as a target for recombinases 12-mer 7 7 D J V1 Molecular explanation of the 12-23 rule Loop of intervening DNA is excised • An appropriate shape can not be formed if two 23-mer flanked elements attempted to join (i.e. the 12-23 rule)
V4 V5 V3 V6 V2 V7 9 V8 V9 9 23-mer 12-mer 7 7 D J V1 CONSEQUENCES OF RECOMBINATION Generation of P-nucleotides
V4 V5 V3 V6 V2 V7 9 V8 V9 9 23-mer 12-mer 7 7 D J V1 Generation of N-nucleotides Terminal deoxynucleotidyl Transferase (TdT) Loop of intervening DNA is excised
A TdT enzim a CD3 régióba illeszt extra-nuklleotid szekvenciákat
Az IgD and IgM típusú B-sejt receptor egyidejű expresszióját alternatív splicing és polyadeniláció teszi lehetővé
How does somatic gene rearrangement (recombination) work? • How is an infinite diversity of specificity generated from finite amounts of DNA? • Combinatorial diversity • 2. How do V region find J regions and why don’t they join to C regions?12-23 rule • How does the DNA break and rejoin? • Imprecisely, with the random removal and addition of nucleotidesto generate sequence diversity • Junctional diversity(P- and N- nucleotides, seeabove)
AZ IMMUNOGLOBULIN SZINTÉZIS FOLYAMATA Szekretált Ig Membrán Ig Golgi ER A H és L láncok külön riboszómákon szintetizálódnak Leader szekvencia Riboszóma mRNS
A B-sejt fejlődés során két ponton történik a génátrendeződés Minőségének ellenőrzése
The pre B-cell receptor monitors the quality of heavy chain rearrangement Mutation in λ5– arrest at Pro-B cell stage SEVERE IMMUNODEFICIENCY Productive µ-chain rearrangement ---assembles pre-BCR Switches off RAG genes, enzymes No further µ-chain rearrangement ALLELIC EXCLUSION Only one specificity
A génátrendetődés sorrendje, kinetikája a B-sejt fejlődés során
Őssejt - HSC Korai pro-B sejt Késői pro-B sejt Nagy pre-B sejt Pre-receptor Perifériás Kis pre-B sejt Éretlen B sejt érett B sejt A B sejt fejlődés szakaszai Csíravonal DH to JH VH to DHJH VHDHJH Y Receptor H+L Az egyes érési fázisokat sejtfelszíni markerek megjelenése (vagy eltűnése) jellemzi
Az autoreaktív receptorral rendelkező B-sejtek a csontvelőben maradnak A B-sejtek begatív szelekciója
A self-reaktív klónokban újra aktiválódik a RAG enzim és a könnyű lánc átrendeződése folytatódik (receptor editing) addig amíg megszűnik a self reaktivitás
Az oldható saját antigénnel reagáló B sejtek (monovalens kötés a sejtfelszíni IgM-hez) funkcionálisan inaktív állapotot (anergiát ) indukál. Ezek az éretleb anergiás sejtek a periférián hamar elpusztulnak IgM monovalens kötés