Download

1 / 36
390 likes | 756 Views
PATHOGENESIS RELATED PROTEINS. INTRODUCTION. Pathogenesis-related proteins, often called PR proteins , are a structurally diverse group of plant proteins that are toxic to invading fungal pathogens.

E N D
INTRODUCTION • Pathogenesis-related proteins, often called PR proteins, are a structurally diverse group of plant proteins that are toxic to invading fungal pathogens. • They are widely distributed in plants in trace amounts, but are produced inmuch • greater concentration in pathogen attack orstress. • Varying types of PR proteins have been isolated from each of several crop plants. Different plant organs, e.g., leaves, seeds, and roots, may produce different sets of PR proteins. • The several groups of PR proteins have been classified according to their function, serological relationship, amino acid sequence, molecular weight, and certain other properties. • PR proteins are either extremely acidic or extremely basic and therefore are highly soluble and reactive.
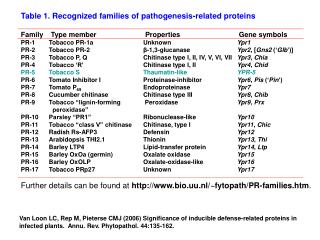
At least 14 families of PR proteins are recognized. The better known PR proteinsare • PR1 proteins (antioomycete andantifungal), • PR2 (b-1,3-glucanases), • PR3(chitinases), • PR4 proteins(antifungal), • PR6 (proteinase inhibitors) • thaumatine-likeproteins, • Defensins, thionins, lysozymes, osmotinlike proteins, lipoxygenases, cysteine-rich proteins, glycine-rich proteins, proteinases, chitosanases, andperoxidases. • There are often numerous isoforms of each PR protein in various host plants. • Although healthy plants may contain trace amounts of several PR proteins, attack by pathogens, treatment with elicitors, wounding, or stress induce transcription of a battery of genes that code for PR proteins.
Occurrence:- 1. In several plant species upon infection with viruses, viroids or bacteria the development of symptoms is accompanied by appearance of one or more new proteins. such protein found in tobacco & recently has been detected in 16species. The occurance of these new proteins is not pathogen specific but determined by the type of reaction in host plant this is indicated that they are hostspecific. All are identified by polyacrylamide gel electrophoresis run either in the absence orin the presence of SDS & stained with general proteinstains. At least tobacco & cowpea ,PRs are found predominantly outside the cytoplasm in the intercellularspaces. They must secreted through plasma lemma.at least four tobacco PRs firstidentified are not glycoproteins nor oxidase or hydrolases such as peroxidases,Ribonuclease, protease & phosphatase which are known to be present in intercellular space. Their resistance to proteases in an obvious requirement for a function inthis environment in which proteolytic enzymeabound.
Properties:- Both tobacco & bean PR elute from gel filtration columns than bulk of protein from healthy leaves i.e it indicate that they are low molecular weight protein & their large polyacrylamide gel electrophoresis due to chargedifferences. They are selectively low pH, where they are soluble than other proteins. Ten PRs can be identified as prominent bands when crude low pH extracts are subjected to the same electrophoreticprocedure. They are highly resistant to the endogenous plant proteases as well as to commercial protease preparation including trypsin.chymotrypsin,papain, & proteasek. They are not protease inhibitor nor produced by treatment of protein extracts fromnon infected leaves withproteases.
Induction of PR protein:- They are induced by plasmolysis. since they are also induced artificially bythe application of a variety of chemicals notably Polyacrylic acid, Benzoic acid derivatives. Various plant hormones as well as culture filtrates from pathogenic fungus or bacterium. It has been proposed that induction might be the result of some type of stress. Ethylene is produced by most plant tissues as a general reaction to stress & PRs can be induced in all tobacco tested to date by ethepon by which ethylene released in plant as well as natural precursor of ethylene i.e 1-aminocyclopropane-1-carboxylic acid (ACC). This gives idea that the induction of PRs is mediated by ethylene, at least in tobacco . 5.Induction under all conditions investigated so far is associated withstimulated ethyleneproduction.
When leaf discs infected with TMV were floated on a solution of amino ethoxyvinylglycine which specially blocks the formation ofACC (1-aminocyclopropane-1-carboxylic acid) in biosynthesis of ethylene. Induction of PRs by ethepon or salicylic acid in tobacco is inhibited by cyclohexamide or D-2-(4-methyl-2,6-dinitroanilin)-N-methyl propionamide both inhibitor of protein synthesis on cytoplasmicribosomes.
Pathogenesis-related (PR) protein1 • Occurance & Molecularnature • The first PR- 1 protein was discovered in1970. • Since then, a number of PR-1 proteins have been identified in • Arabidopsis • Barley • Tobacco • Rice • Pepper • Tomato • Wheat • Maize • These PR-1 having 14 to 17 kD molecular weight and mostly of basicnature. 8
Mode ofaction • Non-expressors of Pathogenesis-Related Genes1 (NPR1) regulate systemicacquired • resistance via regulation pathogenesis related-1 (PR-1) in Arabidopsisthaliana. • The interaction of nucleus-localized NPR1 with TGA transcription factors, after reduction of cysteine residues of NPR 1 by salicylic acid (SA) results in the activation of defense genes of PR-1. In the absence of TAG 2 and/or SA expression of PR-1 not occur in Arabidopsisthaliana. • PR-1 proteins have antifungal activity at the micromolar level against a number of plant pathogenic fungi, including Uromyces fabae, Phytophthora infestans, and Erysiphe graminis. • The exact mode of action of the antifungal activities of these proteins are yet to be identified but a PR-1-like protein, helothermine, from the Mexican banded lizard have been found to be interacting with the membrane-channel proteins of target cells, inhibiting the release of Ca2+. 9
Example:- β-Glucanases Occurance &Properties:- • Plant β-1,3-glucanases (β -1,3-Gs) comprises of large and highly complex genefamilies • involved in pathogen defense as well as a wide range of normal developmentalprocesses. • β -1,3-Gs have molecular mass in the range from 33 to 44 kDa. • These enzymes are found in wide variety of plantslike • Peanut, • Chickpea • Tobacco, etc. and having resistivity against various fungi like Aspergillus parasiticus,A. • flavs, Blumeriagraminis. • These enzymes have wide range of isoelectricpH. • Most of the basic β-1,3-Gs are localized in vacuoles of the plant cells while the acidic β – 1,3-Gs are secreted outside the plantcell. Pathogenesis relatedprotein-2 10
Wounding, hormonal signals like methyl jasmonate and ethylene , pathogen attack like fungus Colletotrichum lagenarium and some fungal elicitors releases frompathogen • cell wall can also induced β-1,3-Gs in the various parts ofplant. • β-1,3-glucanases are involves in hydrolytic cleavage of the 1,3-β-D-glucosidic linkages in β-1,3-glucans, a major componant of fungi cell wall. So that cell lysis and cell death occur as a result of hydrolysis of glucans present in the cell wall offungi. • The enzyme β-1,3-Gs was found to be strongly induced by ultraviolet (UV-B; 280– 320nm) radiation in primary leaves of French bean (Phaseolus vulgaris), so that UV-I induced DNA damage is a primary step for the induction ofβ-1,3-Gs. • β-1,3- glucanases and chitinases are down regulated by combination of auxin and cytokinin while Abscisic acid (ABA) at a concentration of 10 μM markedly inhibited the induction of β-1,3-glucanases but not of chitinases. Mode ofAction 11
Pathogenesis relatedprotein-3 • Occurrence &properties • Example:-Chitinase • Most of Chitinase having molecular mass in the range of 15 kDa and 43kDa. • Chitinase can be isolated from Chickpea, Cucumber,barley. • The main substrate of Chitinases is chitin is a natural homopolymer of β-1,4- linked N- acetylglucosamineresidues. • Chitinases can be divided into two categories: • Exochitinases:- demonstrating activity only for the non-reducing end of the chitinchain. • Endochitinases:- which hydrolyse internal β-1,4- glycosidebonds. • Many plant endochitinases,especially those with a high isoelectric point, exhibit an additional lysozyme or lysozyme likeactivity. • Chitinase and β-1,3-Glucanase are differentially regulated by Wounding, Methyl Jasmonate, Ethylene, andGibberellin. 12
Mode ofaction • Chitinases hydrolyze chitin to soluble oligosaccharides, mainlyN,N-diacetylchitobiose • (G2), which is further hydrolyzed to G1 (N-acetylglucosamine) by GlcNAcases. • In some study, it is also found that Chitinase gene are also expressed in response to stress like cold up to -2 to-5ºC. • These Chitinases have significant antifungal activities against plant pathogenic fungi like • Alternaria sp. For grain discoloration of rice, Bipolaris oryzae for brown spot ofrice. • The mode of action of PR-3 proteins is relatively simple i.e. Chitinases cleaves thecell • wall chitin polymers in situ, resulting in a weakened cell wall and rendering fungalcells • osmoticallysensitive. 13
Chitin BindingProtein • All chitin binding proteins do not possess antifungal activities.CBP can be isolate from plant • sugerbeet, • Hortensia • tobacco • pepper • tomato andpotato • bacteria like Streptomycestendae. • Moleculer weight of the CBP was found to be in the range of 9 kDa to 30 kDa and having basic isoelectricpH. • CBP shows strong inhibitory effect against fungi Aspergillus species, Cercospora beticola, Xanthomonas campestris and many more and several crop fungalpathogen. • Enzymetically CBP has not any function but it binds to insoluble chitin andenhances Pathogenesis relatedprotein-4 hydrolysis of chitin by other enzyme likeChitinase. 16
Mode ofAction • Chitin’s biodegradable and anti-fungal properties are also useful for environmentaland • agriculturaluses. • Chitooligosaccharides, which are the sugar intermediates released during chitin hydrolysis, also are pharmaceuticallyimportant. • CBP, which binds to the insoluble crystalline substrate, leading to structural changes in the substrate and increased substrateaccessibility. • CBP21 strongly promoted hydrolysis of crystalline chitin by chitinases A and C, while it was essential for full degradation by chitinaseB. • CBP variants with single mutations on the largely polar binding surface lost theirability • to promote chitin degradation, while retaining considerable affinity for the polymer. • Thus, binding alone is not sufficient for CBP21 functionality, which seems to depend on specific, mostly polar interactions between the protein and crystallinechitin. 15
Thaumatin likeprotein:- • Occurrence &Properties • Thaumatin-like proteins comprise of polypeptides classes that share homology with thaumatin, sweet protein from (Bennett)Benth. • Thaumatin-like proteins can be isolated from barley, kiwifruit,maize. • Most of the TLPs have a molecular weight in the range of 18 kDa to 25 kDa and have a pH in the range from 4.5 to5.5. • Constitutive levels of Thaumatin-Like Protein is typically absent in healthy plants, with the proteins being induced exclusively in response to wounding or to pathogen attack like Uncinula necator, Phomopsisviticola. • Linusitin is a 25-kDa Thaumatin-like Protein isolated from flaxseeds. • Linustin shows antifungal activity against Alternaria alternata by the mechanismof • membrane permeabilization. Concentration of protein and lipid and composition of cell wall of fungi play a major role in thesemechanisms. Pathogenesis RelatedProtein-5 16
Thaumatin production is induced in katemfe fruit in response to an attack uponthe • plant by viroidpathogens. • Several members of the thaumatin protein family display significant in vitro inhibition of hyphal growth and sporulation by variousfungi. • The thaumatin protein is considered a prototype for a pathogen-response protein domain. This thaumatin domain has been found in species as diverse as rice and Caenorhabditiselegans. • The proteins are involved in systematically acquired resistance and stressresponse • in plants, although their precise role isunknown. • It is induced by attack by viroids, which are single-stranded unencapsulated RNA molecules that do not code forprotein. • The thaumatin protein I consists of a single polypeptide chain of 207residues. • Like other PR proteins, thaumatin is predicted to have a mainly beta structure, witha Mode ofaction high content of beta-turns and littlehelix. 19
Plant proteaseinhibitors:- • The possible role of protease inhibitors (PIs) in plant protection was investigatedas • early as 1947 by, Mickel andStandish. • The term “protease” includes both “endopeptidases” and “exopeptidases” whereas, the term “proteinase” is used to describe only “endopeptidases” (Ryan,1990). • Several non-homologous families of proteinase inhibitors are recognized amongthe • animal, microorganisms and plantkingdom. • Majority of proteinase inhibitors studied in plant kingdom originates fromthree • main families namely leguminosae, solanaceae and gramineae (Richardson,1991). • Many of protease inhibitors are rich in cysteine and lysine, contributing to better and enhanced nutritionalquality. 18
Mode ofaction • These protease inhibitor genes have practical advantages over genes encoding for complex pathways i.e. by transferring single defensive gene from one plant species to another and expressing them from their own wound inducible or constitutive promoters thereby imparting resistance against insect pests (Boulter,1993). • Protease inhibitors also exhibit a very broad spectrum of activity including suppression of pathogenic nematodes like Globodera tabaccum, G. pallida, and Meloidogyne incognita by CpTi (Williamson and Hussey,1996). • These inhibitor families that have been found are specific for each of thefour • mechanistic classes of proteolytic enzymes, and based on the active amino acidin • their “reaction center” (Koiwa et al. 1997), are classified as serine, cysteine, aspartic and metallo-proteases. 19
Ribosome inactivatingprotein • Ribosome-inactivating proteins (RIPs) are toxic N-glycosidases that depurinate the universally conserved sarcin loop of largerRNAs. • This depurination inactivates the ribosome, thereby blocking its further participation in proteinsynthesis. • RIPs are widely distributed among different plant genera and within a varietyof • differenttissues. • Recent work has shown that enzymatic activity of at least some RIPs is not limited to site-specific action on the large rRNAs of ribosomes but extends to depurinationand • even nucleic acid scission of othertargets. • Characterization of the physiological effects of RIPs on mammalian cells has implicated apoptotic pathways. For plants, RIPs have been linked to defense by antiviral, antifungal, and insecticidal properties demonstrated in vitro andin Pathogenesis related Protein (PR-10) transgenicplants. 22
1. Type 1 RIPs, such as pokeweed antiviral protein (PAP), saporin (from soapwort) and barley translation inhibitor, are monomeric enzymes, each with an approximate MW of 30,000. • They are basic proteins that share a number of highly conserved active cleftresidues • and secondary structure within the active site region but are distinctly different in overall sequence homology and posttranslational modifications • 2. Type 2 RIPs, like ricin and abrin, are highly toxic heterodimeric proteins with enzymatic and lectin properties in separate polypeptide subunits, each of approximate MW of 30,000. • One polypeptide with RIP activity (A-chain) is linked to a galactose binding lectin(B- • chain) through a disulfidebond. • 3. Type 3 RIPs, are synthesized as inactive precursors (proRIPs) that require proteolytic processing events to occur between amino acids involved in formation of the active site. They are isolated from maize and barley, although severalclose CLASSIFICATION OFRIP relatives of maize, includingsorghum. 23
Mode ofaction • Type 1 RIPs from Mirabilis expansa roots were active at microgram levels against several soil borne bacterial species, the first such demonstration of antibacterial activity from a plant RIP inbioassays. • In addition, these RIPs were active against a wide variety of both pathogenic and nonpathogenic fungi including Fusarium and Trichodermaspecies. • Type 1 barley RIP was inhibitory to fungal growth on solid media when testedagainst • Trichodermareesei. • Inhibition of growth in liquid media, however, was minimal with barley RIP alonebut • increased dramatically when chitinase was alsoincluded. • A type 2 RIP from seeds of the camphor tree Cinnamomum camphora, was toxic to larvae of mosquito and cottonbollworm. 22
Plant defensins:- • Occurance & Properties • The main groups of antimicrobial peptides found in plants are thionins, defensins and lipid transfer proteins. The name “plant defensin” was coined in 1995 by Terras and collegues. • Plant defensins are small (M.W. 5kDa), basic, cysteine-rich antifungal peptidesranging • from 45 to 54 amino acids, and are positivelycharged. • The first plant defensins were isolated from wheat and barley and were initially classified as a subgroup of the thionin family called theγ-thionins. • since they showed a similar size and the same number of disulfide bridges as α andβ- • thionins. • The plant defensins Rs-AFP1 and Rs-AFP2 from radish, and alfAFP isolated from seeds of the alfalfa plants, are examples of potent antifungalproteins. Pathogenesis relatedprotein-12 23
The structure-activity relationships and modes of action for most of the plant defensins remain unknown. Not all plant defensins have the same mode of action. Some of them exhibit potent antifungal activity in vitro at micromolar concentrations against a broad spectrum of filamentousfungi. • Morphogenic antifungal defensins reduce hyphal elongation and induce hyperbranching, whereas non-morphogenic defensins reduce hyphal elongation without causing any morphologicaldistortions. • The antifungal activity of plant defensins, whether morphogenic or not, is reduced by increasing the ionic strength of the fungal growth assaymedium. • Antifungal plant defensin, Rs-AFP2, appears to act primarily at the cell membraneand • induces rapid Ca2+ uptake and K+ efflux from Neurospora crassa hyphae and it may thus inhibit the growth of filamentous fungi by disrupting cytosolic Ca2+ gradients essential for hyphal tipgrowth. Mode ofaction 24
Example:- • Plant -thionins • Thionins are small, basic plant proteins, 45 to 50 amino acids in length, which include three or four conserved disulfidelinkages. • Thionins are mainly found in seeds where they may act as a defence againstconsumption • by animals. • A barley leaf thionin that is highly toxic to plant pathogens and is involved in the mechanism of plant defence against microbial infections has also beenidentified. • The hydrophobic protein crambin from the Abyssinian kale is also a member ofthe • thioninfamily. Pathogenesis relatedprotein-13 25
Mode ofaction • The proteins are toxic to animal cells, presumably attacking the cell membrane and rendering it permeable, this results in the inhibition of sugar uptake and allows potassium and phosphate ions, proteins, and nucleotides to leak fromcells. • Some thionins are able to inhibit digestive enzymes, bringing thionins to aselect • group of plant proteins synthesized in response toinsect-pests. • Some thionins described until now are capable to inhibit insect α-amylases and others could inhibit serineproteases. • γ –hordothionins isolated from sorghum was the first example of a thionin able to inhibit insect-amylases. 26
Example:- • LIPID-TRANSFER PROTEINS INPLANTS • Properties:- • Lipid-transfer proteins (LTP) are basic, 9-kDa proteins present in highamounts • in higherplants. • LTPs can enhance the in vitro transfer of phospholipids between membranes and can bind acyl chains. • On the basis of these properties, LTPs were participate in membrane biogenesis and regulation of the intracellular fatty acid pools. The principle of the assays is to monitor the transfer of labelled lipids from donor to acceptormembranes. • Acceptor membranes are either natural membranes such asmitochondria, • chloroplasts, plasma membranes, and “microsomes” (endoplasmic rich-fraction) or artificial Membranes (liposomes or lipid vesicles). Donor membranes are either natural membranes orliposomes. • The lipids to be transferred are either radioactive, spin labelled, orfluorescent. • They were found to be secreted and located in the cell wall. Thus, novel roles were suggested for plant LTPs: participation in cutin formation, embryogenesis, defense reactions against phytopathogens, symbiosis, and the adaptation of plants to various environmentalconditions. Pathogenesis relatedprotein-14 27
Their mode of action is not completely understood. A shuttle mechanism has been proposed for the phosphatidylcholine- specific LTP from mammalian cells, which suggests the formation of a phospholipid-LTP complex that interacts with the membrane and exchanges its bound phospholipid with a phospholipid molecule from the membrane. A similar sequence of events has been suggested for plantLTPs. • Such a complex has never been isolated from a plant LTP (and also with nonspecific LTPs from mammalian cells), which suggests that the binding is too weak to allow formation of a stable complex. • In contrast, a strong binding of acyl chains or of lyso-phosphatidylcholine hasbeen • noted for several plantLTPs. • These binding properties are in agreement with the model suggesting that LTPs contain a hydrophobic cavity that can accept one acyl chain but not a whole phospholipidmolecule. Mode ofaction 28
Properties &occurance • Cultured tobacco cells adapted to grow under osmotic stress synthesizeand • accumulate a 26 Kda protein (osmotin) which can constitute as much as 12% • of total cellularprotein. • In cells adapted to NaCl osmotic occurs in twoforms: • An aqueous soluble form (osmotin-I)and • A detergent soluble form (osmotin II) in the approximate ratio of2:3. • Osmotin strongly resembles the sweet protein thaumatin in its molecular weight, amino acid composition, N-terminal sequence, and the presence of a signal peptide on the precursor protein. Thaumatin does not cross-react withantiosmotin. • Immunocytochemical detection of osmotin revealed that osmotin is concentratedin dense inclusion bodies within the vacuole. Although antiosmotin did not label organelles, cell walls, or membranes, osmotin appeared sparsely distributed inthe osmotin cytoplasm. 31
Osmotin and other osmotin-like proteins were shown to have antifungal activity in vitro against a broad range of fungi, including several plantpathogens. • The fungal growth inhibition by osmotin and zeamatin, a maize PR-5protein, • correlated with plasma membrane permeabilization and dissipation ofthe • membrane potential, suggesting a physical interaction between PR-5 proteins and the plasma membrane of sensitive fungi, but the precise mechanism of cytotoxicity remains unknown. • Many of the PR proteins, including osmotin, exhibit clear specificity of their toxicity against fungi, indicating that there must be determinants of sensitivity and resistance in fungal cells osmotin stimulates a mitogen-activated protein kinase (MAPK) signal system in yeast to induce changes in the cell wall that enhance cytotoxicity of this antifungalprotein. • It has been suggested earlier that the cytotoxic action of plant antifungalproteins • could involve activation of signaling cascades, based on the ability of Gprotein Mode ofaction:- inhibitors to block the cytotoxic effect of plantdefensin. 32
OtherProteins • Theis et al, 2003 investigated the inhibitory effects of the antifungal protein (AFP) from Aspergillusgiganteus. • AFP is a highly basic (Isoelectric point 8.8) polypeptide of 51 amino acids with a high content of cysteine, tyrosine, and lysineresidues. • Seetharaman et al. (1996) identified other AFPs, sormatin,, that increase during caryopsis development; they were high at physiological maturity and decrease at combine harvest maturity of thegrain. 31
Mode ofaction • They show that the growth inhibitory effect of the AFP is caused by permebealization of the fungal membranes by using an assay based on the uptake of the fluorescent dye SYTOXGreen. • Pozo et al., 2002 also found the same AFP protein from the Aspergillus giganteus, it promotes charge neutralization and condensation of DNA as demonstrated by electrophoretic mobility shift and ethidium bromide displacementassays. • Hagen et al., 2007 found AFP can inhibit the chitin synthesis by the In situ chitin synthase activityassays. • It indicate that AFP causes cell wall stress and disturbs cell integrity byinactivating • chitin synthase that results in membrane permeability. 32
Function of PR protein:- PRs are synthesized in response to some type of stress, their function analogs tothe one postulated heat shock protein might be protect the plant fromextensive damage. During hypersensitive reaction, PRs are first detectable in a ring around arenecrosis center. In the subsequent phase of slow lesion expansion, they reach the highest concentration at the lesionmargin they also accumulate between lesion in that inoculated leaves & may appear inlower concentration in distant non infectedleaves. the occurrence of PRs merely reflects a particular type of stress which changes the metabolism of the plant & induces as well as resistance further toinfection. 33
Conclusion • PR proteins play important role in disease resistance, seed germination and alsohelp • the plant to adapt to the environmental stress. • The increasing knowledge about the PR proteins gives better idea regarding the development and defense system ofplants. • Primary aspects of the gene regulation of the PR proteins are understood but the study of exact mechanism of gene regulation and receptor cascade will open new ways for the plant genetic engineering technology for cropimprovement. 34
Additional resistance gene(s) against Cladosporium fulvum present on the Cf-9 introgression segment have been shown to be associated with strong PR-protein accumulation (Laugé et al.,1998). In some pathosystems mRNAs for certain PRs members accumulate to similar levels in compatible and incompatible interactions, but the maximum level of expression is reached much faster in the latter (Van Kan et al.,1992). Accumulation of PRs in plants in which resistance is locally or systemicallyinduced. Generalizing this broad research area it can be stated that PRs arerecognized as markers of the systemic acquired resistance (SAR), and PRs genes are involved in the list of the so-called SAR-genes (Ward et al.,1991). Some SAR-inducing chemicals, such as benzothiadiazole (BTH), β-aminobutyric acid (BABA) or 2,6-dichloroisonicotinic acid (DCINA) are harmless commercially supplied compounds and have promising practical application as novel tools inplant Recentstudies protection (Van Loon, 1997; Ku, 2001; Edreva, 2004 and referencestherein). 37
Gene-engineering of PR-5 is another promising strategy for improvement of crop disease resistance, based on the potent plasmolyzing and antifungal effect of this PRs family. Overexpression of a pepper basic pathogenesis related protein 1 gene in tobacco plants enhances resistance to heavy metal and pathogen stresses (Sarowar et al., 2005). For biotechnological purposes PR genes are transferred from novel sources, such as the insectivorous sundew (Drosera rotundifolia L.) (Matušikova et al.,2004). 36