Download

1 / 44
460 likes | 613 Views
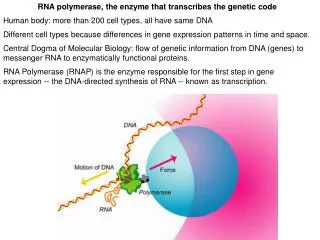
Activities of RNA Polymerase. sequence specific DNA binding -promoters melts DNA to reveal the template strand selects ribonucleotide (not deoxynucleotides) that anneals to template strand polymerizes RNA strand translocates on DNA template, during which it must:

E N D
Activities of RNA Polymerase sequence specific DNA binding -promoters melts DNA to reveal the template strand selects ribonucleotide (not deoxynucleotides) that anneals to template strand polymerizes RNA strand translocates on DNA template, during which it must: -unwind DNA in front of polymerase -unwinds RNA:DNA hybrid -rewinds DNA behind polymerase recognize termination signals in the nascent transcript (or on the DNA template) In addition, the polymerase must be processive (have a high probability of reaching the end of the gene)
Modular Organization of Regulatory Information via Multiple Enhancers stripes of ftz and eve expression in a Drosophila embryo fragments of eve regulatory were inserted upstream of a -Gal reporter and inserted into flies. different regulatory elements gave distinct patterns of -Gal (dark staining) expression. Normal eve expression is shown in red. 1
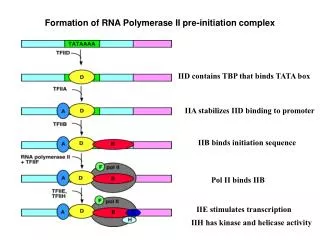
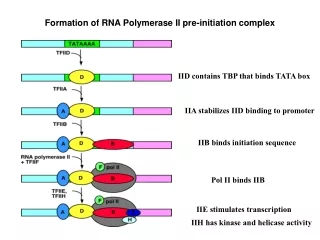
Stages ofTranscription Promoter binding [closed complex] Open complex formation [open complex] Abortive initiation Promoter clearance Elongation
Robert Roeder today collecting sea urchin embryos 1968
Keys to Successful Protein Purification • an abundant source of material • a quantitative assay • must be able determine yield and purity • a strategy for separation • charge • size • hydrophobicity • stability • affinity reagents
Roeder’s Assay, Incorporation of 32P-labeled UTP into RNA he could measure the amount of radioactivity incorporated into RNA (vs that that remained associated with the UTP) 32 note that the phosphate is incorporated into the RNA
Three Different RNA Polymerases in Eukaryotes (Roeder and Rutter, 1969) DEAE-Sephadex functional group on column: polymerase activity (32P incorporation into RNA) protein (measured by UV light absorbance in a spectrophotometer) [salt] (50mM KCl -400 mM)
RNA Polymerases I, II, and III Exhibit Different Sensitivities towards -Amanitin % Maximum activity Amanita phalloides -Amanitin (g/ml)
Eukaryotic Nuclear RNA Polymerases • Enzyme location relative -amanitin • activity sensitivity • Pol I nucleolus 50-70% not inhibited • Pol II nucleoplasm 20-40% inhibited • Pol III nucleoplasm ≈10% species-specific • All three polymerase classes.... • weigh >500,000 D • contain 12-16 subunits • - some conserved across evolution • ’ - like (~200,000 D) • - like (~140,000 D) • - like (~40,000 D) • - some shared among all 3 polymerases • - some unique
Three Classes of Transcription in Eukaryotes RNA polymerase I (pol I) ribosomal RNAs (5.8S, 18S, 28S rRNA) RNA polymerase II (pol II) mRNAs some small nuclear RNAs (snRNAs) non-coding RNAs (mostly of unknown function) RNA polymerase III (pol III) tRNAs 5S RNA some snRNAs small cytoplasmic RNAs (scRNAs)
RNA Polymerase II Underlies the Central Dogma of Molecular Biology Pol I 45S rRNA Pol II Ribosome DNA mRNA Protein Pol III tRNA 5S rRNA this explains our emphasis on the mechanisms of Pol II transcription
Roger Kornberg (1947) (With wife and sons, Stockholm Dec. 2006)
Important things to know about Pol II • The general architecture of the polymerase, including the arrangement of nucleic acids in the active site • That nucleotides likely enter through the “funnel” • That the polymerase is a catalyst that specifically accelerates the rate at which the correctly paired ribonucleotide is added to a growing RNA chain (the exact details of the proposed reaction mechanism are not important to know) • Review, but don’t feel obligated to memorize the exact details of the role of the trigger loop in facilitating catalysis and substrate (I.e. nucleotide) selection • That the polymerase must be able to translocate on the DNA template after it has added a nucleotide to the RNA
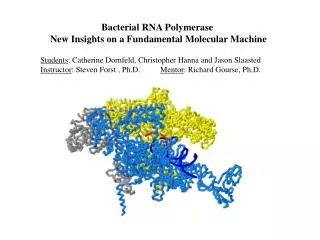
Crystal Structure of Yeast RNA Polymerase II at 2.8 Å Resolution (Cramer et al, 2001)
RNA Pol II (S. cerevisiae) RNAP (T. aquaticus) Rpb2 ’ Rpb1
What is Required for Promoter Function? cis (DNA sequences) vs trans (proteins) identify cis elements by, conservation, mutagenesis and assays of transcription identify trans factors by biochemical (mainly) and genetic (occasionally) approaches
Assays of Promoter Activity in vitro - use a specific promoter - mix with NTPs and extract or purified factors - measure RNA (directly, with 32P-labeled NTPs or indirect assays, see Weaver pp 106-111) in vivo - introducegene of interest into cells transformation (yeast and bacteria) transfection (cultured cells) - fuse promoter to a reporter gene that can be differentiated from normal genes in the recipient cells “reporter gene” - can collect and measure RNA from cells - can measure activity of reporter gene beta-Gal, selectable markers
Identifying Promoter Elements • Deletion mutations • Linker scanning • Point mutations
Class II Promoters Several parts: • Core promoter • Upstream promoter elements • Enhancers, may be far from core promoters gene specific sequence elements found in many core promoters
Core Promoter Elements • In addition to TATA box, core promoters are: • TFIIB recognition element (BRE) • Initiator (Inr) • Downstream promoter element (DPE) • note: the important thing to remember is the TATA box • At least one of the four core elements is missing in most promoters • TATA-less promoters tend to have DPEs • Promoters for highly specialized genes tend to have TATA boxes • Promoters for housekeeping genes tend to lack them
Upstream Elements • Upstream promoter elements are usually found upstream of class II core promoters • Differ from core promoters in binding to relatively gene-specific transcription factors. examples: • GC boxes bind transcription factor Sp1 • CCAAT boxes bind CTF (CCAAT-binding transcription factor) • Enhancers, function in a position and orientation independent manner. (sometimes enhancers are considered to be a distinct type of element)
Identifying the General Transcription Machinery • establish robust in vitro assay with a strong core promoter • AdML adenovirus major late promoter • purify proteins required for transcription • goal is to identify a minimal set of purified proteins with which to reconstitute transcription
What is Required for Promoter Function? Observation: RNA polymerase alone is not capable of accurately initiated, gene-specific transcription. cis (DNA sequences) vs trans (proteins) identify cis elements by, conservation, mutagenesis and assays of transcription -core promoter (esp. TATA box) -upstream elements -enhancers identify trans factors by biochemical (mainly) and genetic (occasionally) approaches -general transcription factors (GTFs) -accessory factors required at all genes transcribed by a polymerase
Identifying the General Transcription Machinery • establish robust in vitro assay with a strong core promoter • AdML adenovirus major late promoter • purify proteins required for transcription • goal is to identify a minimal set of purified proteins with which to reconstitute transcription
RNA Polymerase II Requires Additional Factors for Accurate Transcription Initiation at Promoters (Matsui et al, 1980) Phosphocellulose chromatography
RNA polymerase II Transcription Machinery Number of subunits Pol II 12 GTFs TFIID TFIIB 1 TFIIE 2 TFIIH 9 TFIIF 2 TFIIA 3 Mediator 22 TBP 1 * TAFs 12 *
Proposed Pathway of Initiation Pol II-TFIIF recruited by DNA-D/A/B complex helicases in TFIIH catalyze open complex formation initiation and escape
Mechanism of Initiation of RNA Polymerase II Transcription Pol Pol H TBP H F BN BC BC TBP Factor Role TBP Promoter recognition; configures DNA to the pol II surface TFIIB Promoter recognition; pol II recruitment; directs DNA path, stabilizes early transcribing complex; coupling of RNA synthesis to promoter clearance TFIIE Recognizes closed complex, recruits TFIIH TFIIF Captures nontemplate strand upon melting TFIIH Untwisting of promoter DNA (helicase), CTD phosphorylation (kinase)
Transcription Elongation • ...is slow compared to DNA replication • 20-40 nucleotides / second • Typical 1° transcript is ≈20,000 nts., corresponds to ≈10 minutes / transcript. Long transcripts can take hours to complete • … is regulated • TFIIF suppresses pausing • TFIIS rescues arrested complexes • others • …. polymerases stalled at the 5’ ends of genes appear to be common • ...may involve proof-reading (observed in vitro) • ...is coupled to DNA repair
Activating domains can be replaced by randomly selected sequences (Ma and Ptashne, 1986) beta-gal activity Q. what are the essential features of activation domains? Approach: random e. coli sequences cloned downstream of Gal4 DBD and expressed in yeast containing a -gal reporter with Gal4 sites in its promoter 16
Activating domains can be replaced by randomly selected sequences (Ma and Ptashne, 1986) • random e. coli sequences cloned downstream of the Gal4 DBD and expressed in a yeast strain containing a -gal reporter with Gal4 sites in its promoter • ~1% of all the clones activated transcription • activating sequences did not resemble known proteins, no catalytic domains etc. -activation domains unlikely to have enzymatic activity 4. negatively charged residues common 17
HIS3 +1 +12 -Gal core UASH/core Overexpression of Gal4 from the strong ADH promoter inhibits promoters that lack Gal4 binding sites (Gill and Ptashne, 1988) decreased expression of reporter genes lacking Gal4 binding sites when Gal4 levels are high UASG= GAL enhancer (binds Gal4) UASH= HIS3 enhancer UASC= CYC1 enhancer 18
Activator Interference or ‘Squelching’ activator B activator A UAS TATA box 19
Activator Interference or ‘Squelching’ activator B activator A UAS TATA box hypothesis? 20
What is the Limiting Target of Activators? Eukaryotic activators do not bind to RNA pol II polymerase and therefore do not directly recruit polymerase to promoters. Activators may, however, indirectly recruit RNA polymerase by recruiting factors (often called co-activators) that serve as a physical bridge between activator and polymerase. ‘TFIID hypothesis’ ‘Holoenzyme hypothesis’ 21
ChIP (Chromatin Immunoprecipitation) TBP Formaldehyde crosslink TBP Shear chromatin by sonication TBP Immunoprecipitation TBP Reverse crosslinks, PCR input control: reverse crosslinks and analyze sample prior to IP 22
Extent of TBP binding correlates with promoter activity (Li et al, 1999) TBP crosslinks to active promoters 23
RNA Polymerase II Transcription Machinery Number of subunits Pol II 12 GTFs TFIID TFIIB 1 TFIIE 2 TFIIH 9 TFIIF 2 TFIIA 3 Mediator 22 TBP 1 * TAFs 12 * 24
The ‘TFIID Hypothesis’ 1. TAFs provide surfaces for the interaction of TFIID with activators. 2. TFIID recruits polymerase in vitro assays suggest specific activator-TAF contacts predictions? 25
Activated Transcription in the Absence of TAFIIs Oelgeschlager et al., 1998 western blot demonstrating depletion of TAFIIs see p97 of Weaver or p769 of Watson for a description of the Western blot technique in vitro transcription shows that - transcription is abolished in the TFIID depleted extract - TBP is sufficient to restore activated transction - 4 different activators were tested 26 no transcription after depletion of TFIID and TAFs