Download

1 / 33
350 likes | 587 Views
Invertebrate nerves and nervous systems. Peter Shaw. A lot to cover in 1 lecture!. Aim for today: 1: basics of nervous function (anatomy + signal transmission) 2: Anatomy of invertebrate nervous systems 3: a few sense organs . What are nervous systems?

E N D
Invertebrate nerves and nervous systems Peter Shaw
A lot to cover in 1 lecture! Aim for today: 1: basics of nervous function (anatomy + signal transmission) 2: Anatomy of invertebrate nervous systems 3: a few sense organs
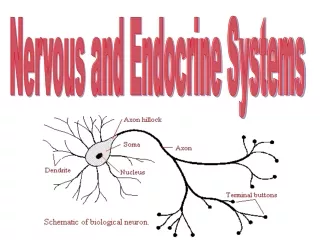
What are nervous systems? They are networks of excitable cells that process information, generally resulting in bodily action. Input: information from sense organs; output: muscular action. Nervous systems are found in all metazoa, and only in metazoa. Sponges lack them, although at a biochemical level all the basic features of nerve cells can be found in parazoan cells. The basic unit of a nervous system is a nerve cell, called a neuron. (Sometimes, I think incorrectly, spelled neurone). Nerve cells are invariably long, since they are involved in communication/transmission. They are usually 10-20um diameter.
And the real thing. Idealised neuron (is vertebrate – ignore the collar like structures and see later..)
It is most important to realise that neurons do not function like wires carrying electricity. A much better analogy would be with transistors relaying a pulse of current. Neurons fire by carrying a depolarising signal. This needs explaining: All cells have a bounding membrane, the plasma membrane, which isolates them from their immediate environment. Even inside a healthy body the composition of bulk body fluids (blood plasma, lymph etc) is quite different to the chemistry inside cells. Indeed, one of the many ways to kill a cell is to puncture its plasma membrane; many antibiotics and natural toxins work this way.
The chemical gradient across a cell membrane is manifested in many ways, one of which is an imbalance in cation composition. Intracellular fluid (cytoplasm) is depleted in calcium and sodium, but elevated in potassium, when compared to extra-cellular fluids. (Calcium ions act as a label of extra-cellular invasion, and are present at very low concentrations intra-cellularly, probably because of their ability to immobilise phosphorus). Cytoplasm is also elevated in organic acids wrt blood. This ionic gradient is maintained by metabolic expenditure: energy is continuously spent pumping sodium out of cells, and potassium inside.
This ionic imbalance generates a charge gradient across cell membranes: the inside of a cell is negative with respect to the outside, by roughly 30 mV. They are said to be polarised, and the 30mv is called the resting potential. This is normal and healthy; all cells are always negatively charged, with one exception. The exception concerns neurons. Neurons contain special proteins in their cell walls which act as ionic gates; they are able to open briefly to destroy the membrane’s insulation. (Note: This is transient; a prolonged opening of the gates would kill the cell).
Neurons have ‘gates’ in their membranes which, when opened, allow entry of sodium, and others which allow exit of potassium. These gates are voltage-sensitive: a crucial part of the mechanism.
When the gates are activated, there is a rapid re-distribution of cations (caused purely by diffusion), which reverses the cell’s polarisation: for a few milliseconds the affected region of a cell’s membrane becomes positively charged on the inside wrt the outside. This phenomenon is known as depolarisation. The sequence is that the sodium gates are opened first, causing an inrush of sodium, changing the membrane potential from 30mv negative to 40ish mv positive. This transient spike of altered polarity is the action potential.
Once depolarised, the membrane-bound potassium gates open, while the sodium shut again. Potassium diffuses out, the sodium pumps continue their work, and within a few more milliseconds normal polarity is re-established. Thus sections of a neuron’s membrane can be in 1 of 2 states: resting potential, or action potential. This is roughly equivalent to the 1/0 logical states used in computing (BUT don’t take the analogy too far). Note that this depolarisation is local on the cell membrane, with only a small region affected at anyone time (unlike a transistor, where the entire silicon surface behaving the same way).
Transmission within cells Within cells it is transmitted via the effect of the depolarisation on membrane gates. The immediate effect of depolarisation is to change the electrical field in the surrounding membrane, and this electrical field affects the membrane-bound proteins, causing the cation gates to open. Thus a depolarised region causes adjacent regions also to depolarise. Once the cation gates have opened, they undergo a brief refractory period when they no longer respond to the electrical field. The effect of this is to ensure that the wave of depolarisation is uni-directional: it cannot go backwards because the region behind an advancing front is recovering from depolarisation.
Depolarisation normally occurs at one end of a neuron (the dendrites), and progresses along over the soma to the axon, where a wave of depolarisation spreads along to the far end of the cell, which can be as much as 10m off in extreme cases.
Transmission between cells Between cells, nerves transmit information chemically. At the end of an axon there is NO physical contact with adjacent neurons. The region where one nerve transmits to another is called the synapse. Between nerve cells there is always a gap, the synaptic cleft, in the order of 100 nm. Vacuoles with neurotransmitter Pre-synaptic membrane Post-synaptic membrane With receptors for transmitters
At the end of an axon is a specialised section of membrane (the pre-synaptic membrane), holding vesicles containing specialised chemicals called neurotransmitters. Each neuron will generally only hold one neurotransmitter. The best known is called acetylcholine, which is also the transmitter by which nerves stimulate muscles. (The neuro-muscular junction behaves in exactly the same way as the nerve-nerve synapse described below).
When an action potential reaches the pre-synaptic membrane, some vesicles fuse with the cell membrane and release molecules of neurotransmitter into the synaptic cleft. These molecules rapidly (0.5ms ish) diffuse across the tiny space of the synaptic cleft, and encounter the post-synaptic membrane of the recipient dendrite. This membrane has special receptors which bind the neurotransmitter, and having bound to the transmitter they briefly open a sodium gate. This causes a local depolarisation. If enough depolarisation occurs, the dendrite experiences an action potential, which is then propagated down the dendrite, over the soma and down the axon: one nerve has transmitted its signal to another.
Transmission between cells contd. This mechanism allows the output from several nerve cells to be added together: if 2 axons both converge onto one dendrite, a low stimulation from either neuron may be inadequate to trigger an action potential, whereas both axons firing together could cause a big enough depolarisation for an action potential to occur. A form of addition! Similarly, subtraction can be emulated by one neuron releasing a neurotransmitter than causes increased polarisation (by opening potassium gates)
Finally, there remains the problem that neurotransmitter will remain in the synaptic cleft for long periods (several seconds), during which time it will continue to stimulate the post-synaptic membrane. To deal with this, synaptic clefts always contain mechanisms to rapidly remove neuro-transmitter. This can be enzymatic (acetylcholine), or membrane-bound pumps in the pre-synaptic membrane (most CNS transmitters).
The speed of conduction. Clearly the faster information travels the better. It turns out that speed of conduction varies with the size of the cell; roughly speed is proportional to the square root of cell diameter. Some invertebrates need to coordinate rapid movements over large distances; they have to use “giant” axons. In the common squid Loligo, contraction of the mantle for swimming depends on a giant axon, 0.5mm diameter. Usefully, this is big enough to stick needles into, and the cell remains active for hours after removal from the animal. Most of our understanding of basic neurophysiology has come from this giant axon. Sabellid polychaetes also have giant fibres innervating fan retractor muscles.
Vertebrates have improved on this basic system by wrapping neurons on an insulating sheath of myelin (Swann cells), forcing the depolarisation to move by jumping between successive gaps in the insulation (nodes of Ranvier). This results both in higher conduction speeds, and in speed increasing linearly with axon diameter. A few invertebrates have independently evolved something similar, though apparently not as effectively as vertebrates.
Vertebrate neurons have a myelin sheath of Schwann cells, forcing the signal to jump from one node of Ranvier to the next.
General function of nervous systems: To collect and process information about the surrounding environment, then to produce appropriate muscular actions. General trend: A progression from diffuse nerve nets to highly centralised systems with differentiated sensory organs The simplest examples are found in the cnidaria:
The lowest level of organisation is found in the cnidaria, where the rather short nerve cells form a diffuse net around the body. These animals have a rather limited behavioural repertoire: retract tentacles into mouth when food is caught, pulsate the bell by contraction of a circular muscle (medusoid forms). Actually some cnidaria have a few more sophisticated manoeuvres. Here are a few, but this list is not exhaustive: 1: the basal muscle of anemones allows very slow lateral movement (“walking”). 2: Adjacent anemones “fight” by release of nematocysts from specialised organs (acontia) around the mouth. 3: Solitary free-living corals (Fungia) can right themselves if overturned. 4: Physalia can dip its gas bladder in water to keep the bladder wall moist.
The cnidarian senses that we know of are touch and chemosensitivity (allowing synchronisation of gamete release). Nematocysts fire without nervous command - the release is a local action by the individual cell. One small radiation of especially dangerous species (the box jellies cubomedusae) have light-sensitive organs (a complex eye including a lens) at each corner of the bell. These do not seem to connect to any obvious nerve mass, but these animals seem able to avoid objects, and engage in complex manoeuvres such as swimming through the supports of a pier without touching. Chironex fleckeri and its eye
Echinoderms have a superficially similar, simple nervous system. They have a nerve ring (pentagonal of course) around the mouth, with branches running off down each arm. Again, these animals have a very limited behavioural repertoire (based on chemical senses) and slow movements with much “local” coordination of tube feet - ie no great need for a complex central controller. All other metazoa have a centralised nervous system with a differentiated central nerve cord and some form of cephalic ganglion (“brain”). Touch, chemical senses and vision are ubiquitous (though vision is often secondarily lost).
Cnidarian – nerve net Flatworms, annelids and arthropods have paired ventral nerve cords running from a “brain” (ganglion) around the oesophagus
The commonest pattern among invertebrates is for the central nervous system to run ventrally along the axis of the body, terminating in a large cephalic ganglion running around the oesophagus (the circumoesophogeal ganglion). This is found clearly in flatworms, and all protostomic phyla (annelids, and arthropods + other phyla). There is a general principle that the development of the nervous system correlates with lifestyle; the faster an animal moves, the more coordination is needed to survive or capture food, the more developed will be its nervous system.
It is tempting to assume that all animals are like us and “think” with the cephalic ganglia. In fact many actions in many invertebrates are coordinated by their closest ganglion, with the “brain” sending higher level commands such as targeting food or avoiding danger, while the muscles coordinating legs / body wall muscles work autonomously. (We have one vestige of this: the burn reflex. When a limb touches something hot, the signal for retraction is sent by the spinal cord, and retraction has started by the time you are consciously aware of the pain.)
Sense organs Another huge topic, which I can barely scrape the surface of today. Sense organs convert environmental information into nervous signals; I’ll just mention 2 standard invert senses today; balance and sight.
Statoliths and statocysts These are balance organs, found widely throughout the metazoa. They involve a highly sensitive organ lined with hair cells, which transmit a signal when the hairs are bent. They get bent by a hard dense object, typically a sand grain or a calcareous lump (statolith). One of the nicest experiments in classical zoology is to replace the statolith in a lobster’s statocyst with an iron filing. The lobster behaved normally. Then hold a magnet over the lobster and it tries to walk upside down.
Vision The sense of sight is one of the animal kingdom’s biggest breakthroughs, giving very accurate spatial information over immense distances (especially out of water). There are a wide variety of apparently different organs used here. Note in particular the dichotomy between unitary eyes (vertebrates, cephalopods) and compound eyes (arthropods). Compound eyes turn out to be excellent for local movement detection, but very inefficient at providing the high resolution of vertebrate eyes.
Just before you consider yourself to be superior to a fly on the basis of your high resolution eye, ponder this: The gene that turns on eye development in mammals (ie YOU) has been expressed in the leg of a fruit fly, where the fly grew a perfect compound eye. Your eye and theirs have a common ancestor. The complete data from the fly eye experiments appeared in Science, Halder et al., v.267, pp 1788-1792, 3/24/95. The Pax -6 gene from a mouse (= human aniridia gene = Drosophila eyeless gene) was expressed in flies; one fly had 14 eyes growing in various odd parts of its body!!