Download

1 / 24
240 likes | 400 Views
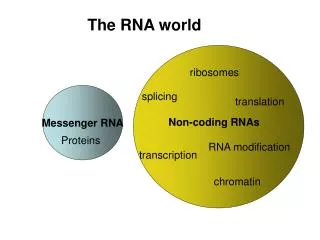
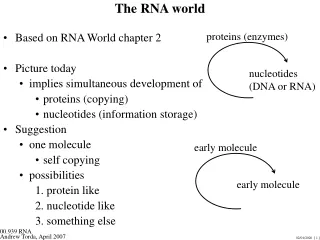
V6 RNA world. short name full name function mRNA, rRNA, tRNA, you know them well ... snRNA small nuclear RNA splicing and other functions snoRNA small nucleolar RNA nucleotide modification of RNAs Long ncRNA Long noncoding RNA various miRNA MicroRNA gene regulation

E N D
V6 RNA world short name full name function mRNA, rRNA, tRNA, you know them well ... snRNA small nuclear RNA splicing and other functions snoRNA small nucleolar RNA nucleotide modification of RNAs Long ncRNA Long noncoding RNA various miRNA MicroRNA gene regulation siRNA small interfering RNA gene regulation SS 2009 – lecture 6 Biological Sequence Analysis 1
RNA structure Single stranded RNA molecules frequently require a specific tertiary structure. The scaffold for this structure is provided by secondary structural elements which are H-bonds within the molecule. This leads to several recognizable "domains" of secondary structure like hairpin loops, bulges and internal loops. RNA hairpin 2RLU Stem loop 1NZ1 www.rcsb.org SS 2009 – lecture 6 Biological Sequence Analysis 2
miRNAs microRNAs (miRNA) are single-stranded RNA molecules of 21-23 nucleotides in length, which regulate gene expression. miRNAs are encoded by DNA but not translated into protein (non-coding RNA). Instead, each primary transcript (a pri-miRNA) is processed into a short stem-loop structure called a pre-miRNA and finally into a functional miRNA. Mature miRNA molecules are partially complementary to one or more messenger RNA (mRNA) molecules. Their main function is to down-regulate gene expression of this mRNA. www.wikipedia.org SS 2009 – lecture 6 Biological Sequence Analysis 3
snRNAs • Small nuclear RNA (snRNA) are found within the nucleus of eukaryotic cells. • They are transcribed by RNA polymerase II or RNA polymerase III and are involved in a variety of important processes such as • RNA splicing, • regulation of transcription factors or RNA polymerase II, and • maintaining the telomeres. • They are always associated with specific proteins. The complexes are referred to as small nuclear ribonucleoproteins (snRNP) or sometimes as snurps. • A large group of snRNAs are known as small nucleolar RNAs (snoRNAs). • These are small RNA molecules that play an essential role in RNA biogenesis and guide chemical modifications of rRNAs, tRNAs and snRNAs. • They are located in the nucleolus and the cajal bodies of eukaryotic cells. www.wikipedia.org SS 2009 – lecture 6 Biological Sequence Analysis 4
Long ncRNAs Long noncoding RNAs (long ncRNAs) are non-protein coding transcripts longer than 200 nucleotides. This limit is due to practical considerations including the separation of RNAs in common experimental protocols. Additionally, this limit distinguishes long ncRNAs from small regulatory RNAs such as miRNAs, short interfering RNAs (siRNAs), Piwi-interacting RNAs (piRNAs), snoRNAs etc. www.wikipedia.org SS 2009 – lecture 6 Biological Sequence Analysis 5
XIST One prominent well-understood ncRNA is the ncRNA XIST that silences the inactive X chromosome by the ncRNA XIST. To normalize the copy number of X chromosomes between male and female cells, transcription of XIST RNA from one of the two female X chromosome is involved in recruiting Polycomb group proteins (PcG) to trimethylate histone H3 on lysine 27 (H3K27me3), rendering the chromosome transcriptionally silent. (wait for second half of this lecture). PcGs thereby remodel chromatin. Polycomb Repressive Complex 2 (PRC2) is a PcG that is comprised of H3K27 histone methyl transferase (HMTase) EZH2 and core components Suz12 and EED, apparently initiates this histone modification. Subsequently, Polycomb Repressive Complex 1 (PRC1) is another PcG that maintains this modification and promotes chromatin compaction. Plath, Science 300, 131 (2003) SS 2009 – lecture 6 Biological Sequence Analysis 6
Hotair: another important ncRNA Because many HMTase complexes lack DNA-binding domains but possess RNA-binding motifs, it has been postulated that ncRNAs may guide specific histone modification activities to discrete chromatin loci. Rinn et al. showed that HOTAIR ncRNA binds PRC2 and is required for robust H3K27 trimethylation and transcriptional silencing of the HOXD locus. Hox genes are a group of related genes that specify the anterior-posterior axis and segment identity of metazoan organisms during early embryonic development. These genes are critical for the proper number and placement of embryonic segment structures (such as legs, antennae, and eyes). HOTAIR may therefore be one of the long-sought-after RNAs that interface the Polycomb complex with target chromatin. Rinn, Cell 129, 1311(2007) SS 2009 – lecture 6 Biological Sequence Analysis 7
Conservation of Hotair HOTAIR is expressed in posterior and distal sites, indicating the conservation of anatomic expression pattern from development to adulthood. Interestingly, this transcript has very high nucleotide conservation in vertebrates (99.5%, 95%, 90%, and 85% sequence identity in chimp, macaque, mouse, and dog genomes, respectively). Yet it contains many stop codons with little amino acid sequence conservation amongst vertebrates. These results suggest that HOTAIR may function as a long ncRNA. Rinn, Cell 129, 1311(2007) SS 2009 – lecture 6 Biological Sequence Analysis 8
Hotair mechanism A potentially attractive model of epigenetic control is the programming of active or silencing histone modifications by specific noncoding RNAs. Just as transcription of certain ncRNA can facilitate H3K4 methylation and activate transcription of the downstream Hox genes, distant transcription of other ncRNAs may target the H3K27 HMTase PRC2 to specific genomic sites, leading to silencing of transcription and establishment of facultative heterochromatin. Model of long ncRNA regulation of chromatin domains via histone-modification enzymes. Transcription of ncRNAs in cis may increase the accessibility of TrxG proteins such as the histone methyl transferase ASH1 or MLL or directly recruit them, leading to H3K4 methylation and transcriptional activation of the downstream HOX gene(s). In contrast, recruitment of PRC2 is programmed by ncRNAs produced in trans, which targets PRC2 activity by as-yet-incompletely-defined mechanisms to target loci. PRC2 recruitment leads to H3K27 methylation and transcriptional silencing of neighboring HOX genes. Rinn, Cell 129, 1311(2007) SS 2009 – lecture 6 Biological Sequence Analysis 9
What is epigenetics? Epigenetics refers to alternate phenotypic states that are not based in differences in genotype, and are potentially reversible, but are generally stably maintained during cell division. Examples: imprinting, twins, cancer vs. normal cells, differentiation, ... The narrow interpretation of this concept is that of stable differential states of gene expression. A much more expanded view of epigenetics has recently emerged in which multiple mechanisms interact to collectively establish - alternate states of chromatin structure, - histone modification, - associated protein composition, - transcriptional activity, and - in mammals, cytosine-5 DNA methylation at CpG dinucleotides. Laird, Hum Mol Gen 14, R65 (2005)
Basic principles of epigenetics:DNA methylation and histone modfications The human genome contains 23 000 genes that must be expressed in specific cells at precise times. Cells manage gene expression by wrapping DNA around clusters (octamers) of globular histone proteins to form nucleosomes. These nucleosomes of DNA and histones are organized into chromatin, the building block of a chromosome. Rodenhiser, Mann, CMAJ 174, 341 (2006) Bock, Lengauer, Bioinformatics 24, 1 (2008)
Epigenetic modifications Strands of DNA are wrapped around histone octamers, forming nucleosomes. These nucleosomes are organized into chromatin, the building block of a chromosome. Reversible and site-specific histone modifications occur at multiple sites through acetylation, methylation and phosphorylation. DNA methylation occurs at 5-position of cytosine residues within CpG pairs in a reaction catalyzed by DNA methyltransferases (DNMTs). Together, these modifications provide a unique epigenetic signature that regulates chromatin organization and gene expression. Rodenhiser, Mann, CMAJ 174, 341 (2006)
Cytosine methylation 3-6 % of all cytosines are methylated in human DNA. How many cytosines are in „normal“ DNA? How many CpG islands are in „normal“ DNA? Mammalian genomes contain much less (only 20-25 %) of the CpG dinucleotide than is expected by the G+C content. This is typically explained in the following way: As most CpGs serve as targets of DNA methyltransferases, they are usually methylated. 5-Methylcytosine, whose occurrence is almost completely restricted to CpG dinucleotides, can easily deaminate to thymine. If this mutation is not repaired, the affected CpG is permanently converted to TpG (or CpA if the transition occurs on the reverse DNA strand). Hence, methylCpGs represent mutational hot spots in the genome. If such mutations occur in the germ line, they become heritable. A constant loss of CpGs over thousands of generations can explain the scarcity of this special dinucleotide in the genomes of human and mouse. Esteller, Nat. Rev. Gen. 8, 286 (2007)
effects in chromatin organization affect gene expression Schematic of the reversible changes in chromatin organization that influence gene expression: genes are expressed (switched on) when the chromatin is open (active), and they are inactivated (switched off) when the chromatin is condensed (silent). White circles = unmethylated cytosines; red circles = methylated cytosines. Rodenhiser, Mann, CMAJ 174, 341 (2006)
Basic principles of epigenetics:DNA methylation and histone modfications Changes to the structure of chromatin influence gene expression: genes are inactivated (switched off) when the chromatin is condensed (silent), and they are expressed (switched on) when chromatin is open (active). These dynamic chromatin states are controlled by reversible epigenetic patterns of DNA methylation and histone modifications. Interestingly, repetitive genomic sequences are heavily methylated, which means transcriptionally silenced. Enzymes involved in this process include - DNA methyltransferases (DNMTs), - histone deacetylases (HDACs), - histone acetylases, - histone methyltransferases and the - methyl-binding domain protein MECP2. Rodenhiser, Mann, CMAJ 174, 341 (2006)
DNA methylation Typically, unmethylated clusters of CpG pairs are located in tissue-specific genes and in essential housekeeping genes, which are involved in routine maintenance roles and are expressed in most tissues. These clusters, or CpG islands, are targets for proteins that bind to unmethylated CpGs and initiate gene transcription. In contrast, methylated CpGs are generally associated with silent DNA, can block methylation-sensitive proteins and can be easily mutated. The loss of normal DNA methylation patterns is the best understood epigenetic cause of disease. In animal experiments, the removal of genes that encode DNMTs is lethal; in humans, overexpression of these enzymes has been linked to a variety of cancers. Rodenhiser, Mann, CMAJ 174, 341 (2006)
Uptake of methyl groups DNA methylation patterns fluctuate in response to changes in diet, inherited genetic polymorphisms and exposures to environmental chemicals. Methyl groups are acquired through the diet and are donated to DNA through the folate and methionine pathways. Consequently, changes in DNA methylation may occur as a result of low dietary levels of folate, methionine or selenium. This can lead to diseases such as neural tube defects, cancer and atherosclerosis. Imbalances in dietary nutrients can lead to hypomethylation (which contributes to improper gene expression) and genetic instability (chromosome rearrangements). E.g. hyperhomocysteinemia and global hypomethylation have been observed in vitro in atherosclerosis models. In advanced stages of atherosclerosis, hyperproliferation may further contribute to DNA hypomethylation and altered gene expression. Rodenhiser, Mann, CMAJ 174, 341 (2006)
Epigenetic regulation during development Surani, Hayashi, Hajkova, Cell 128, 747 (2007)
Epigenetic regulation during development Surani, Hayashi, Hajkova, Cell 128, 747 (2007)
Epigenetic regulation during development Surani, Hayashi, Hajkova, Cell 128, 747 (2007)
Epigenetic signals in ES cells and in differentiated cells Bernstein, Meissner, Lander, Cell 128, 669 (2007)
Large scale identification of functional ncRNA Eric Lander, Whitehead Institute, MIT John Rinn, Harvard Medical School Gringeras, Nat. Biotech. 27, 346 (2009) Commentary on Guttman article SS 2009 – lecture 6 Biological Sequence Analysis 24