Download

1 / 40
430 likes | 712 Views
Signal transduction = conversion of a signal from one form into another. extracellular signals are converted into different biochemical forms, distributed to different sites in the cell, and sustained and amplified (cascade) as it proceeds

E N D
Signal transduction = conversion of a signal from one form into another. • extracellular signals are converted into different biochemical forms, distributed to different sites in the cell, and sustained and amplified (cascade) as it proceeds • result of intracellular signaling may be changes in the cytoskeleton and secretory apparatus, as in the release of secretory vesicles • final destination may be the nucleus, where activation of transcription factors turns on new gene expression and cell division and differentiation may be induced.
Lymphocytes combine both “neuronal and contact-dependent. See Table 1
Each cell responds to a limited but complex set of signals and each signal can induce different responses in different target cells. • A typical cell is exposed to thousands of different signal molecules, but responds selectively only to certain signals, depending on which receptors it expresses. (Same DNA, selected gene expression in individual cell types) • Each cell responds to the same signal in different ways. • One signal, binding to one type of receptor protein, can cause a multitude of effects in the target cell - shape, movement, metabolism, exocytosis of stored granules, gene expression • Several molecular signals, binding to their receptors, can amplify or modify each other’s responses.
The same signal molecule can induce different responses in different target cells. Different cell types express different types of receptors, which generate quite different intracellular signals. And different intracellular signaling pathways.
A cell depends on many different extracellular signals to survive, divide, differentiate, and even to die when necessary. Programmed cell death = apoptosis
The extracellular signal is passed on from the receptor to an intracellular signaling molecule. Then from one intracellular signaling molecule to another in a signaling cascade. Which 1. physically transfers the signal to the cell machinery that will make the response. 2. transforms the signal into a molecular form able to stimulate the response 3. amplifies the signal, making it stronger. 4. Distributes the signal to different effector mechanisms. 5. And allows other intracellular signals to interface - modifying the end result.
A receptor protein can be located in the plasma membrane. It transduces an extracellular signal into an intracellular signal, initiating a signaling cascade that relays the signal into the cell, amplifying and distributing it. Many of the steps in the cascade can be modified by other signaling events.
Receptors can be located inside the cell. In this case the signaling molecule must be able to diffuse through the cell membrane - small hydrophobic signal molecules. These receptors can be located in the cytosol or the nucleus.
The best known hydrophobic signal molecules are steroid hormones - see Table 1.
Cortosal binds a receptor in the cytosol. The complex is then transported into the nucleus via the nuclear where it binds to specific regulatory sequences in the DNA and activates gene transcription.
There are three main classes of cell-surface receptors which generate different intracellular signals. The flow of ions produces an electrical effect. An activated G-protein subunit is released to initiate the intracellular signaling cascade. An enzyme activity is switched on at the cytoplasmic end, which generates a variety of intracellular signals.
G-protein Ion channel Same extracellular signal - different receptors - different intracellular signals generated lead to different results.
Foreign substances, drugs and poisons, and pathogens bind to cell-surface receptors. • Heroin, nicotine, tranquilizers … • mimic the natural ligand for the receptor • block or over stimulate the receptor • HIV, polio virus… • invade the cell and replicate
Ion channels convert chemical signals into electrical ones • Serve for rapid transmission • important in neuron neurotransmitter release across synapses and muscle cell contraction • important in many cell types in opening calcium channels
Intracellular signaling molecules act as molecular switchs - turning the next signaling molecule on (active).
G-protein receptors form the largest family of cell-surface receptors. They mediate responses to an enormous diversity of extracelluar signal molecule, including hormones and neurotransmitters. Given their tremendous variety, they all have a similar structure, consisting of a singly polypeptide chain that transverses the membrane seven times - a seven-pass transmembrane receptor. Ligand-binding domain G protein binding damain
The receptor undergoes a conformational change when the signaling molecule binds, enabling the cytoplasmic domain to bind to the inactive G-protein.
When the receptor binds the inactive (GDP)G-protein, it is activated. The alpha subunit ejects its GDP and replace it with GTP, detaches from the beta/gamma subunits. Both are now activated and both interact with target molecules in the plasma membrane α
These subunits remain activated a short time, interact with other molecules, activating or inactivating them. Finally, GTP is hydrolized, and the alpha subunit reassociates with the beta/gamma subunit. Cholera toxin enters epithelial cells of the gut, modifies the alpha subunit so that it remains active. Shut off is as important as turn on. Na+ flows out continually - water follows
Targets for G-protein subunits are either ion channels or membrane-bound enzymes. Interaction or an ion channel with a G-protein immediately changes the behavior of the cell. The flow of ions effects other molecules, like water in cholera, immediately. This flow in ions can also effect the complex intracellular signaling pathways - Ca+ - see Fig 23.
Interactions of G subunits with membrane-bound enzymes have more complex consequences, by starting a cascade of intracellular signaling molecules including cAMP and PIP, kinases, etc.
The most frequent target molecules for G proteins are adenylate cyclase, which in turn produce the small signaling molecule cyclic AMP (cAMP) and phospholipase C (PLC), which produces inositol trisphosphate (IP3) and diacylglycerol (DAG). The coupling may be either stimulatory or inhibitory (inhibitory G proteins). Stimulatory G proteins lead to the activation of a cascade of second messengers, which now move away from the membrane to the to the target effect
Increase in cyclic AMP in response to an extracellular signal Blue is a low level of cAMP, yellow is medium, and red is high.
Example: preformed vesicles fuse with the plasma membrane and insulin is secreted.
Activation of gene transcription by a rise in cAMP concentration. cAMP-dependent protein kinase (A-Kinase) phosphorylates a cascade of kinases and other signaling molecules Table 2 - in different cells different sets of intracellular signaling molecules are available - leading to different effects
G-protein-linked receptors activate another signaling pathway via phospholipase C, which gives rise to an increase in cytosolic Ca++. High cytosolic calcium levels trigger Ca++ dependant enzymes like protein kinase C and calmodulin. See Table 3 for some responses
Ca++ influx into the cytosol triggers many biological effects. The normal concentration of free Ca++ is kept low by membrane pumps. This results in a steep electrochemical gradient of Ca++ across ER and plasma membrane (chapter 12). When a signaal transiently opens Ca++ channels Ca++ rushes into the cytosol, triggering changes in Ca++-dependent proteins. The inositol phospholipid signaling pathway occurs in almost all eucaryotic cells and affects many different target proteins - Ca++-binding proteins. Calmodulin is present in all eucaryotic cells. Binds a wide range of targets, including CaM-Kinases.
Many different stimulatory and inhibitory G-proteins exist, creating complex signaling patterns specific to each receptor and each target cell. The effects here are simplified for clarity
Amplification in the light-induced cascade in rod photoreceptor cells. Remember, although not shown in these simplified diagrams, other signals modulate each step and each step has intrinsic off switches. This is equally as important as “on switches”.
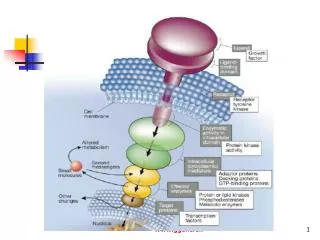
Enzyme-linked Receptors include growth factor receptors which signal complex cellular reactions leading to proliferation, differentiation, or survival. Responses to growth factors are usually slow (hours). Enzyme-linked receptors also mediate direct, rapid effectos on the cytoskeleton, controlling cell movement and shape (phagocytosis). Abnormalities in signaling via enzyme-linked receptorscan lead to cancer. The largest class of enzyme-linked receptors contain a cytoplasmic domain which functions as a tyrosine protein kinase.
These receptors usually have only one transmembrane segment (α helix). Signal molecules must cross-link, bring together more than one receptor in order to induce a signal. No allosteric “shape changes”. Once brought together the cytoplasmic domains (kinases) phosphorylate each other.
Phosphorylation triggers the assembly of many intracellular signaling molecules which can bind to the newly phosphorylated sites on the enzyme-linked receptor’s cytoplasmic tail. Each of these intracellular signaling molecules can act coordinately to induce many biochemical changes which inturn trigger a complex response.
Tyrosine phosphatases remove phosphates, turning off the signal. Or recpetors are endocytosed and destroyed in lysosomes. Turning off an intracellular signal is as important as turning it on. Figure 29 depicts activation of Ras which, when mutated, is a pathway to cancer. A proto-oncogene.
Acetylcholine acts at a G-protein-linked receptor on heart muscle to make the heart beat more slowly by the effect of the G protein on a K+ channel. Which one or more of the following would enhance this effect of acetylcholine? A. A high concentration of a non-hydrolyzable analog of GTP. B. Modification of the acetylcholine-receptor-linked G-protein a subunit by cholera toxin. C. Mutations in the acetylcholine receptor that weaken the interaction between the receptor and acetylcholine. D. Mutations in the acetylcholine receptor that weaken the interaction between the receptor and the G protein. E. Mutations in the G-protein b subunit that weaken the interaction between ab and the K+ channel.