Download

1 / 35
360 likes | 541 Views
E. coli Aspartate Transcarbamoylase (ATCase), Cont’d. Van Vliet et al. (1991) Proc Natl Acad Sci USA 88, 9180-3. Sakash et al. (2000) Prot Sci 9, 53-63. West et al. (2004) J Biol Chem 279, 945-51. Van Vliet et al. (1991) Proc Natl Acad Sci USA 88, 9180-3.

E N D
E. coliAspartate Transcarbamoylase(ATCase), Cont’d Van Vliet et al. (1991) Proc Natl Acad Sci USA 88, 9180-3. Sakash et al. (2000) Prot Sci 9, 53-63. West et al. (2004) J Biol Chem 279, 945-51.
Van Vliet et al. (1991) Proc Natl Acad Sci USA 88, 9180-3. • Sakash et al. (2000) Prot Sci 9, 53-63. • West et al. (2004) J Biol Chem 279, 945-51. The use of nucleotide analogs to evaluate the mechanism of the heterotropic response of Escherichia coli aspartate transcarbamolylase
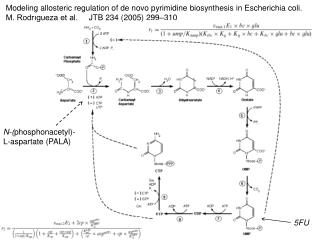
E. coli ATCase Carbamoyl-phosphate (CP) + L-aspartate --> N-carbamoyl-L-aspartate Homotropic ligand: L-aspartate (positive cooperativity) Homotropic regulation fits to MWC model; High-affinity R state and low-affinity T state Heterotropic ligands: ATP (allosteric activator) UTP and CTP (allosteric inhibitor) Heterotropic regulation does not fit as well to MWC; Mechanism????
E. coli ATCase Holoenzyme viewed along 3-fold axis. Binding mode of bisubstrate analog PALA to active site. The allosteric sites are located at the distal ends of the R-chains, 60 Å away from the nearest active site, and bind each effector.
Regulatory nucleotides bind to same site in allosteric (AL) domain of R-chain
Regulation of ATCase by nucleotides-- summary of results from structural and site-directed mutagenesis studies of R-chain. • His20r and Lys94r interact with triphosphate portion of nucleotide and are important for effector binding and resulting heterotropic response. • Triphosphate form of nucleotides are necessary for high binding affinity and for full regulatory response. • Lys60 interacts with 2’-OH of nucleotides and with O2 carbonyl of CTP… plays important role in CTP binding and inhibition and in discrimination between CTP and ATP. • Base portion of nucleotides interacts with backbone carbonyl of Tyr89r and with backbone carbonyl and amine of Ile12r.
Importance of interactions between either ribose or base in nucleotide effector with aa residues in nucleotide binding site investigated using nucleotide analogs: Also 2’-deoxy, 3’-deoxy, or 2’,3’-dideoxy versions tested
Experiments carried out at saturating [CP] and at low [Asp] = 0.5 x [Asp]0.5… conditions in which heterotropic effects are relatively large
Complex response of ATCase to Me-ATP and Me-PTP Very high affinity behavior at low [Me-NTP] gives way to normal affinity binding at high [Me-NTP]
Effects of other base changes on ATCase heterotropic response Me-GTP 150 11.2 Me-GTP No effect Experiments carried out at saturating [CP] and at low [Asp] = 0.5 x [Asp]0.5… conditions in which heterotropic effects are relatively large
Response of ATCase K60rA mutant to nucleotide effectors • Lys60r interacts with 2’OH and O2 carbonyl of CTP. • K60rA mutant is not inhibited by CTP, but is activated by ATP similar to w.t. ATCase. • Although GTP and ITP inhibit w.t. enzyme, neither has any effect on the K60rA mutant. • This suggests that GTP and ITP inhibit w.t. ATCase by a mechanism similar to CTP.
Deoxy forms of ATP activate ATCase as well as (or better than) ATP itself Affinity for 2’-dATP is 2-fold higher than ATP. Affinities for 3’-dATP and ddATP are much lower than ATP.
Deoxy forms of CTP inhibit ATCase as well as CTP itself Affinity for 2’-dCTP is 3-fold higher than CTP. Affinity for and ddCTP is 15-fold lower than CTP.
Interactions between nucleotide effectors and regulatory binding sites are mediated by hydrogen bonding plus hydrophobic contacts and electrostatic attraction.
Interactions between the backbone of ATCase and the external amine groups of ATP and CTP are important for proper orientation of nucleotide in the binding site. • Contacts between backbone carbonyls of Ile12r and Tyr89r and N6 of adenine are important for nucleotide orientation. • Replacement of N6-NH2 with -H, -Cl, or -SH still allows enzyme activation, though not as well as natural ATP effector. • These analogs and ATP have similar electrostatic surface potentials, so electrostatic interactions with enzyme should be similar. • Ile12r backbone interactions with nucleotide are important not only for effector binding, but also for transmission of the regulatory signal.
Regulatory nucleotides bind to same site in allosteric (AL) domain of R-chain
Ile12r as allosteric transmitter: • van der Waals interaction between Ile12r side chain and backbone of Glu62r. • Glu62r is part of S3’ b-strand on which Arg55r, Lys56r, and Lys60r reside, all of which are implicated in heterotropic mechanism. • Backbone displacement of Ile12r upon nucleotide binding may be propagated to Glu62r, causing movement of S3’ b-strand.
Ile12r/Tyr89r interactions with CTP/ATP are similar, and may act as pivot point for orientation of nucleotides. • Lys60r seems to be the most important residue for discrimination between ATP and CTP. • Lys60r side chain may be pushed one way to interact with N3 of ATP and another way to interact with O2 carbonyl of CTP. • Movement of S3’ b-strand within 10-strand antiparallel b-sheet that spans regulatory subunit may dictate overall conformation of nucleotide binding site. • Binding of ATP or CTP may induce local conf. changes in b-sheet which are propagated throughout the enzyme, resulting in activation/inhibition. • Movement of S3’ b-strand resulting from interaction of nucleotides with Lys60r or Ile12r may influence conformation of b-sheet, allowing ATP and CTP to stabilize their respective conformational states of ATCase.
Interaction between O6 carbonyl of CTP and Lys60r is important for enzyme inhibition. • K60rA mutant not inhibited by CTP, affinity for CTP down 100x. • Specific contact with O2, not 2’-OH of CTP important for binding, discrimination, inhibition. • Inhibition of ATCase by purine nucleotides GTP, ITP, XTP consistent with these nucleotides binding in a syn conformation. • Negative surface potential around N7 and O6 carbonyl resemble that around N3 and O2 carbonyl of CTP/UTP. • Me-GTP alters electrostatic surface potential around N7 --> activation, not inhibition. • ATP and purine nucleotides that activate ATCase have surface potentials that resemble each other, but not CTP or purine nucleotides that inhibit ATCase. • Complementary electrostatic interactions between nucleotide and effector binding site are important for binding and orientation, thereby inducing either activation or inhibition.
Upshot: ATCase regulatory site does not discriminate between purines and pyrimidines based on ring size. Complementary electrostatic interactions are the main thing.
Multiple stages in activation of ATCase by Me-PTP and Me-ATP • 6 regulatory binding sites of ATCase are non-equivalent. • CTP and ATP bind to 3 sites with high affinity and 3 sites with low affinity. • Low affinity sites may actually be UTP binding sites that allow UTP to cause additional inhibition of ATCase only in the presence of CTP. • Unusual response of enzyme to Me-PTP and Me-ATP may reflect these analogs binding to both types of sites. (However this doesn’t explain the 3rd phase, where activation decreases). • Alternatively, Me-PTP and Me-ATP might bind in both syn and anti conformations, one with high and the other with low affinity. • Cooperative effects?
Van Vliet et al. (1991) Proc Natl Acad Sci USA 88, 9180-3. • Sakash et al. (2000) Prot Sci 9, 53-63. • West et al. (2004) J Biol Chem 279, 945-51. Heterotropic interactions in aspartate transcarbamolylase: Turning allosteric ATP activation into inhibition as a consequence of a single tyrosine to phenylalanine mutation
Hydrophobic interactions between AL and Zn domains of R-chain: Tyr77r is inserted into a hydrophobic pocket comprised of Leu32r and Leu76r from AL, plus Val106r and Leu151r from Zn domain.
Y77rF Mutant: • Loss of homotropic cooperativity for Asp. • 3x higher apparent Km for Asp. • 2x lower apparent Vmax.
ATP acts as an inhibitor, not an activator of Y77rF ATCase. Inhibition resembles that of CTP.
Y77rF Summary: • Mutation abolishes homotropic cooperativity between catalytic subunits, but does not abolish heterotropic effects. • Therefore homo- and hetero-tropic effects uncoupled. • Hydrophobic core between AL and Zn domains of R-chain, together with R1C4 and C1C4 interfaces, are important for establishment of T state and consequently for homotropic cooperativity between catalytic sites. • Normal sensitivity to CTP but altered sensitivity to ATP indicates Y77 interactions with hydrophobic pocket are not essential for transmission of CTP signal, but are important for ATP signalling. • Inversion of ATP signal indicates that interface controls how the ATP signal is interpreted by the enzyme.
E. coli ATCase Holoenzyme viewed along 3-fold axis. Binding mode of bisubstrate analog PALA to active site. The allosteric sites are located at the distal ends of the R-chains, 60 Å away from the nearest active site, and bind each effector.
Van Vliet et al. (1991) Proc Natl Acad Sci USA 88, 9180-3. • Sakash et al. (2000) Prot Sci 9, 53-63. • West et al. (2004) J Biol Chem 279, 945-51. A fluorescent probe-labeled Escherichia coli aspartate transcarbamoylase that monitors the allosteric conformational state
T state R state
Ribbon diagram of the three-dimensional structure of the C1 and C4 catalytic chains of aspartate transcarbamoylase, with a model of Cys-241 labeled with pyrene, in the T state (A) and R state (B) pyrene excimer West, J. M. et al. J. Biol. Chem. 2004;279:945-951
Pyrene fluorescence: • fluoresces from excited monomer state. • also fluoresces from an excited-state dimer, or “excimer”. • results from specific interaction between excited monomer and ground-state monomer. • “molecular ruler”, range 0-20 Å, perfect for studies of many protein conformational changes. • attached to a mutant ATCase (C47A/A241C) in the 240’s loop region, the pyrene excimer becomes a sensitive probe for the allosteric transition of ATCase.
Fluorescence emission spectra for the PALA saturation of the C47A/A241C-pyrene enzyme Results obtained with mutant ATCase holoenzyme, but not with mutant C-chain alone, demonstrating that pyrene fluorescence change is a function of the overall allosteric confromational change. West, J. M. et al. J. Biol. Chem. 2004;279:945-951
Aspartate saturation curves for the wild-type holoenzyme and the C47A/A241C-pyrene enzyme WT West, J. M. et al. J. Biol. Chem. 2004;279:945-951
Influence of ATP (circles) and CTP (squares) on the activity (filled symbols) and relative fluorescence coefficient (open symbols) of the C47A/A241C-pyrene enzyme West, J. M. et al. J. Biol. Chem. 2004;279:945-951