Download

1 / 43
510 likes | 1.41k Views
513341 Biochemistry I Chapter 7 ENZYME KINETICS. Dr. PORNTIP CHAIMANEE Chemistry Department Faculty of Science Silpakorn University. Enzymatic Analysis. Enzyme (E). E + S P + E. Time. Temperature. Substrate (S). Product (P).

E N D
513341 Biochemistry I Chapter 7 ENZYME KINETICS Dr. PORNTIP CHAIMANEE Chemistry Department Faculty of Science Silpakorn University
Enzymatic Analysis Enzyme (E) E + S P + E Time Temperature Substrate (S) Product (P) อัตราเร็วของปฎิกิริยา(velocity, v) = d[P]/dt หรือ -d[S]/dt
Principle of spectrophotometry L Detector Monochromator I I0 Absorption recorder Light source Sample Absorption A = -log I/Io = e.c.L Beer-Lambert law
Assay of enzyme activity Time Absorbance (sec) of product 15 0.075 30 0.15 45 0.20 60 0.26 75 0.32 90 0.36 105 0.40 120 0.44 150 0.51 180 0.57 210 0.62 240 0.66 E Absorption Time (sec)
AP Reaction velocity, rate of reaction, v v = -d[A ]/dt =d[P]/dt v = k[A] k, rate constant (rate coefficient, specific reaction rate)
VelocityorRatedetermination Initial rate (slope at t = 0) [Product] Concentration Velocity (vO) = slope at t = t1 [Substrate] t = t1 t = 0 Time
1902 Adrian Brown Experiment Sucrose + H2O glucose + fructose yeast enzyme (invertase) Reaction rate become independent to [Sucrose], when [Sucrose] >>> [Invertase enzyme]
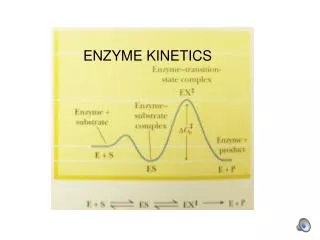
INITIAL RATE, STEADY STATE CONDITIONS k1k2 k-1 E + S ES P + E E = free enzyme ES = enzyme-substrate complex k1, k-1,k2 : rate constants [S] >>> [E] + [ES]
k1 k2 k-1 E + S ES E + P 1 2 1 Presteady state conditions, a few milliseconds 2 Steady state conditions : a few minutes
Michaelis-Menten Kinetics at steady state assumptiond [ES]/dt = 0 = k1[E][S] –k-1[ES] – k2[ES] solve for [ES] [ES] = [E][S] k1/(k-1 + k2) Define KM (Michealis Constant) KM = (k-1 + k2)/k1 then [ES] = [E][S]/KM rearrange to give KM = [E][S]/[ES]
Michaelis-Menten Kinetics KM = [E][S]/[ES] from [E] = [E]T- [ES] KM = ([E]T- [ES] )[S] / [ES] KM[ES] = ([E]T - [ES])[S] [ES] = [S] ( [E]T - [ES] ) / KM [ES] = [E]T [S] / KM+ [S] Multiply both side by k2 k2[ES] = k2[E]T [S] / KM+ [S]
Michaelis-Menten Kinetics =kcat[E]T Maximum velocity If vo = Vmax / 2 Then KM= [S] When KM = [S] , The reaction runs at half maximum velocity
Michaelis-Menten Kinetics Vmax =k2[E]T =kcat[E]T kcat = Vmax [E]T kcat(turnover number) = maximum number of substrate turn into product in one second by one enzyme molecule (one active site) vO=Vmax [S] KM +[S] at physiological condition [S] <<< KM = Vmax KM [S] = (kcat/KM) [E]T[S] Specificity constant Howrapidlyanenzymecan workatlow [S]
Michaelis-Menten Kinetics E + S ES P + E k1 k2 k-1 kcat/KM = k2 / KM = k1k2 / k-1+k2 k2 k-1 >>> kcat/KM = k1 k 1 not more than diffusion controlled limit ~ 108-109 M-1 sec-1 if kcat/KM ~ 108-109 M-1 sec-1 so that enzyme reach the maximum enzymatic catalytic efficiency
Michaelis-Menten Kinetics Significance of KM • KM = [E][S]/[ES] and KM = (k-1 + k2)/k1. • KM is the apparent dissociation constant of the ES complex. A dissociation constant (KD) is the reciprocal of the equilibrium constant (KD=KA-1). KM is a measure of a substrate’s affinity for the enzyme (but it is the reciprocal of the affinity). • If k1,k-1>>k2, the KM=KD. • KM is the substrate concentration required to reach half-maximal velocity (vmax/2). A small KM means the sustrate binds tightly to the enzyme and saturates (max’s out) the enzyme. • The microscopic meaning of Km depends on the details of the mechanism.
Michaelis-Menten Kinetics The significance of kcat • vmax = kcat Etot • kcat: For the simplest possible mechanism, where ES is the only intermediate, and dissociation is fast, then kcat=k2, the first order rate constant for the catalytic step. • If dissociation is slow then the dissociation rate constant also contributes to kcat. • If one catalytic step is much slower than all the others (and than the dissociation step), than the rate constant for that step is approximately equal to to kcat. • kcat is the “turnover number”: indicates the rate at which the enzyme turns over, i.e., how many substrate molecules one catalytic site converts to substrate per second. • If there are multiple catalytic steps (see trypsin) then each of those rate constants contributes to kcat. • The microscopic meaning of kcat depends on the details of the mechanism.
Michaelis-Menten Kinetics Significance of kcat/KM • kcat/KM is the catalytic efficiency. It is used to rank enzymes. A big kcat/KM means that an enzyme binds tightly to a substrate (small KM), with a fast reaction of the ES complex. • kcat/KM is an apparent second order rate constant v=kcat/KM[E]0[S] • kcat/KM can be used to estimate the reaction velocity from the total enzyme concentration ([E]0). kcat/KM =109 => diffusion control. • kcat/KM is the specificity constant. It is used to distinguish and describe various substrates.
The values of kcat ,KMand(kcat/KM) for some enzymes and substrates
Variationofinitialrate (v0) as a functionof [S]withconstantconcentrationofenzyme
Variation of initial rate(v0) as a function of [S] with constant concentration of enzyme The initial velocity approaches a maximum at high [S].
Data analysis: The Lineweaver-Burk plot (1934) Michaelis-Menten Equation y = m.x + b
Lineweaver-Burk-Plot y intercept = 1/vmax x intercept = -1/KM the slope is KM/vmax
ENZYME INHIBITION การทำงานของเอนไซม์อาจถูก ยับยั้งหรือรบกวน โดยสารบางชนิดที่สามารถ ยื้อแย่ง กับ substrate ในการเข้าจับกับเอนไซม์ทำให้ประสิทธิภาพ ในการทำงานลดลงหรือเสียไป สารบางชนิดดังกล่าวจะเรียกว่า สารยับยั้ง หรือ inhibitor TWO TYPES OF INHIBITORS 1. IRREVERSIBLE INHIBITORS 2. REVERSIBLE INHIBITORS
IRREVERSIBLE INHIBITORSCompounds Reactive group of enzymes Examples CN- enzymemetalions(eg. Fe, Zn ,Cu ) Cytochrome oxidase Sarin -SHgroupofcysteineAcetylcholineesterase Parathion -SHgroupof cysteineAcetylcholineesterase DIPF(Nervegas) -OHgroupofserineAcetylcholineesterase O 2(CH3)-CH-O-P-O-CH-(CH3)2 F Chymotrypsin Iodoacetamide -SHgroupofcysteineGlyceraldehyde-3- I-CH2-C-NH2 phosphatedehydrogenase O
Nerve Poisons Inactivated by paraoxonase Used by terrorists in Tokyo subway, 1995 Inhibited Acetylcholinesterase used as insecticides DIPF so toxic that it has been used as military nerve gas.
Penicillin is an irreversible of the enzyme glycoprotein peptidase glycoprotein peptidase
REVERSIBLE INHIBITORS k1 k2 k-1 E + S ES E + P General principle: a) Any inhibitor that displaces the equilibrium E + S ES modifies the Km b) Any inhibitor that changes the maximum concentration of ES modifies the Vmax
Competitive Inhibitors Substrate Enzyme k1 k2 k-1 E + S ES P + E + I EI + S NO REACTION P ES complex KI Competitive inhibitor Inactive enz EI complex
Variation of initial rate(v0) as a function of [S]with and without competitive inhibitor Vmax Uninhibited v0 Competitive inhibition Vmax/2 KM aKM [S]
Competitive Inhibitors Vmax[S] KM (1+[I]/KI) +[S] = 1/vo = KM ( 1 + [I]/KI ) . 1/ [S] + 1/ Vmax
A double-reciprocal (Lineweaver-Burk) plot with and without competitive inhibitor KM increased Vmaxunchanged KM increased Vmaxunchanged Increasing [I] 1/vo E =Succinate dehydrogenase S =Succinate -2OC-CH2-CH2-CO2- I =Malonate -2OC-CH2-CO2- Uninhibited -1/KM 1/[S] -1/aKM
Mixed (competitive and uncompetitive) Inhibition Substrate Enz k1 k2 k-1 P E + S ES P + E + + I I EI + S ESI NO REACTION ES complex Noncompetitive inhibitor KI K/ I Inactive enz
Variation of initial rate(v0) as a function of [S]with and without noncompetitive inhibitor Vmax Uninhibited v0 Vmax/2 Noncompetitive inhibition KM [S]
A double-reciprocal (Lineweaver-Burk) plot Of Mixed (competitive and uncompetitive) Inhibitor KM changed Vmaxchanged Increasing [I] 1/vo 1/aVmax Uninhibited 1/Vmax -1/KM 1/[S]
Uncompetitive Inhibitors Substrate Enz k1 k2 k-1 E + S ES P + E + I ESI NO REACTION P ES complex K/ I Uncompetitive inhibitor Inactive enz
Variation of initial rate(v0) as a function of [S]with and without uncompetitive inhibitor Uninhibited v0 Uncompetitive inhibition aKM KM [S]
A double-reciprocal (Lineweaver-Burk) plot with and without uncompetitive inhibitor KM diminished Vmaxdiminished Slope= KM /Vmax 1/vo Increasing [I] 1/aVmax Uninhibited 1/Vmax -1/KM 1/[S] -1/aKM Chemistry Science Silpakorn University Dr. Porntip Chaimanee
HIV PROTEASE INHIBITORS -Phe-Pro HIV protease substrate Saquinavir Hoffman-LaRoche KI=0.40 nM Ritonavir KI=0.015 nM Abbott Laboratories
X-Ray structure of HIV-1 protease (a) Uncomplexed inhibitor (b) in complexed with its inhibitor saquinavir
Effect of temperature on velocity vo Arrhenius / Law Thermal denaturation Optimum temperature Temperature
Effect of pH on velocity Pepsin vo Trypsin pH 1.5 7.7 Optimum pH