Download

1 / 75
790 likes | 1.19k Views
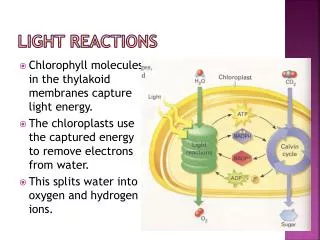
Light Reactions of Photosynthesis. 2 H 2 O + 2 NADP + + 8 photons → O 2 + 2 NADPH + 2 H +. ANIMATION. Integration of photosystems I and II in chloroplasts. The "Z scheme“ evolved by combining 2 bacterial RCs. Resembles that of green sSulfur bacteria. Resembles that of purple bacteria.

E N D
Light Reactions of Photosynthesis 2 H2O + 2 NADP+ + 8 photons → O2 + 2 NADPH + 2 H+ ANIMATION
Integration of photosystems I and II in chloroplasts. The "Z scheme“ evolved by combining 2 bacterial RCs. Resembles that of green sSulfur bacteria Resembles that of purple bacteria. Reaction center chlorophylls lie close to the exiton acceptor preventing internal conversion (fluorescence). This fixed orientation mimics the solid state. analogous to cyt c
Cyclic vs. Nonclyclic PS • animation
All electron carriers are bound to a nearly symmetric dimer. Participants are positioned like the bacterial RC in purple bacteria. Photosystem II of the cyanobacterium Synechococcus elongates
Light Reactions of Photosynthesis PS II 4 P680 + 4 H+ + 2 PQB + 4 photons → 4 P680+ + 2 PQBH2
The supramolecular complex of PSI and its associated antenna chlorophylls
Light Reactions of Photosynthesis PS I 4 P700 + 2 H+ + 2 NADP+ + 4 photons → 4 P700+ + 2 NADPH
Electron and proton flow through the cytochrome b6f complex Plastoquinol (PQH2) formed in PSII is oxidized by the cytochrome b6f complex in a series of steps like those of the Q cycle in the cytochrome Complex III of mitochondria. One electron from PQH2 passes to the Fe-S center of the Rieske protein, the other to heme bL of cytochrome b6. The net effect is passage of electrons from PQH2 to the soluble protein plastocyanin, which carries them to PSI. f for frons Heme groups
Non cyclic electron flow (PSI + PS II) produces a proton gradient + NADPH • Cyclic electron flow (PSI) produces a proton gradient only • Calvin cycle requires ATP and NADPH in a ratio of 3:2 • The 2 PS are physically separated to prevent exitons from leaving P680 and transferring to P700. Why would this happen? • (longer wavelength, lower energy)
Localization of PSI and PSII in thylakoid membranes. PSII is present almost exclusively in the appressed regions (granal lamellae [stacks], in which several membranes are in contact) , and PSI almost exclusively in nonappressed (stromal lamellae) regions, exposed to the stroma. LHCII is the "adhesive" that holds appressed lamellae together.
Balancing of electron flow in PSI and PSII by state transition noncyclic cyclic Accumulation of plastoquinol stimulates a protein kinase that phosphorylates a Thr residue in the hydrophobic domain of LHCII, which reduces its affinity for the neighboring thylakoid membrane and converts appressed regions to nonappressed regions (state 2). A specific protein phosphatase reverses this regulatory phosphorylation when the [PQ]/[PQH2] ratio increases.
H+ released into lumen. Water-splitting activity of the oxygen-evolving complex Water splitting complex passes 4 electrons, 1 at a time, back to P680+. The electrons lost from the multinuclear Mn center pass one at a time to an oxidized Tyr residue in a PSII protein, then to P680+
Light-Induced Redox Reactions and Electron Transfer Cause Acidification of Lumen Because the volume of the lumen is small, a few hydrogen ions dramatically change the pH: lumen = pH 5, stroma pH = 8!!! The proton-motive force across the thylakoid membrane drives the synthesis of ATP… sound familiar?? Thylakoid membrane is impermeable to hydrogen ions. Reaction centers, electron carriers and ATP synthases are located in this membrane Uncouplers decouple light absorption from ATP synthesis.
In vitro ATP synthesis • Incubate chloroplasts in pH 4 buffer in the dark. Buffer slowly entered thylakoids lowering the pH to 4. • Add ADP and Pi and suddenly increase the pH to 8. (How would you do this?) IN THE DARK. • !!!! ATP produced!!!
Flow of Protons: Mitochondria, Chloroplasts, Bacteria According to endosymbiotic theory, mitochondria and chloroplasts arose from entrapped bacteria Bacterial cytosol became mitochondrial matrix and chloroplast stroma
Comparison of the topology of proton movement and ATP synthase orientation in the membranes of mitochondria, chloroplasts, and the bacterium E. coli.
Dual roles of cytochrome b6f and cytochrome c6 in cyanobacteria reflect evolutionary origins. Cyanobacteria use cytochrome b6f, cytochrome c6, and plastoquinone for both oxidative phosphorylation and photophosphorylation.
CHAPTER 20Carbohydrate Biosynthesis in Plants and Bacteria CO2 assimilation in photosynthetic organisms Photorespiration in C3 plants Avoiding photorespiration in C4 plants Key topics:
Introduction to Anabolic Pathways Anabolism: how to build biomolecules Plants are extremely versatile in biosynthesis Can build organic compounds from CO2 Can use energy of sunlight to support biosynthesis Can adopt to a variety of environmental situations
Plant versatility • Autotrophic • Nonmobile/motile • CHO synthesis occurs in plastids • Plants synthesize thick cell walls exterior to the cell containing the bulk of the cell’s CHO—how do they do this???
Plants and Photosynthetic Microorganisms Support the Life of Animals and Fungi Plants capture the energy from the ultimate energy source and make it available via carbohydrates to animals and fungi
Photosynthetic organisms use the energy of sunlight to manufacture glucose and other organic products, which heterotrophic cells use as energy and carbon sources.
Biological reproduction occurs with near-perfect fidelity (although no 2 zebras have exactly the same stripes!). Zebras are herivores.
CO2 Assimilation Occurs in Plastids Self-reproducing organelles found in plants and algae Enclosed by a double membrane Have their own small genome Most plastid proteins are encoded in the nuclear DNA The inner membrane is impermeable to ions such as H+, and to polar and charged molecules
Amyloplasts are pastids without the internal membrane or pigments. Amyloplasts filled with starch (dark granules) are stained with iodine in this section of Ranunculus (buttercups)root cells.
Origin and Differentiation of Plastids Plastids were acquired during evolution by early eukaryotes via endosymbiosis of photosynthetic cyanobacteria Plastids reproduce asexually via binary fission The undifferentiated protoplastids in plants can differentiate into several types, each with a distinct function Chloroplasts for photosynthesis Amyloplasts for starch storage Chromoplasts for pigment storage Elaioplasts for lipid storage Proteinoplasts for protein storage
internal membranes lost Proplastids in nonphotosynthetic tissues (such as root) give rise to amyloplasts, which contain large quantities of starch. All plant cells have plastids, and these organelles are the site of other important processes, including the synthesis of essential amino acids, thiamine, pyridoxal phosphate, flavins, and vitamins A, C, E, and K.
CO2 Assimilation The assimilation of carbon dioxide occurs in the stroma of chloroplasts via a cyclic process known as the Calvin cycle The key intermediate, ribulose 1,5-bisphosphate is constantly regenerated using energy of ATP The key enzyme, ribulose 1,5-bisphosphate carboxylase / oxygenase (Rubisco), is probably the most abundant protein on Earth The net result is the reduction of CO2 with NADPH that was generated in the light reactions of photosynthesis
Early studies of the Calvin cycle • Design an experiment to discover the pathway for carbon assimilation • First intermediate recognized was 3-PGA • Search for a 2 carbon acceptor---FAILURE • Actual acceptor….
CO2 Assimilation The assimilation of carbon dioxide occurs in the stroma of chloroplasts via a cyclic process known as the Calvin cycle The key intermediate, ribulose 1,5-bisphosphate is constantly regenerated using energy of ATP The key enzyme, ribulose 1,5-bisphosphate carboxylase / oxygenase (Rubisco), is probably the most abundant protein on Earth The net result is the reduction of CO2 with NADPH that was generated in the light reactions of photosynthesis
The Structure and Function of Rubisco Rubisco is a large Mg++-containing enzyme that makes a new carbon-carbon bond using CO2 as a substrate
Structure of ribulose 1,5-bisphosphate carboxylase (rubisco). Ribbon model of form II rubisco from the bacterium Rhodospirillum rubrum. The subunits are in gray and blue. A Lys residue at the active site that is carboxylated to a carbamate in the active enzyme is shown in red. The substrate, ribulose 1,5-bisphosphate, is yellow; Mg2+ is green.
Mg2+ is coordinated in a roughly octahedral complex with six oxygen atoms: one oxygen in the carbamate on Lys201; two in the carboxyl groups of Glu204 and Asp203; two at C-2 and C-3 of the substrate, ribulose 1,5-bisphosphate; and one in the other substrate, CO2. Central role of Mg2+ in the catalytic mechanism of rubisco.
First stage of CO2 assimilation: rubisco's carboxylase activity.
Catalytic Role of Mg++ in Rubisco’s Carboxylase Activity • Notice that Mg++ is held by negatively charged side chains of • glutamate, • aspartate, and • carbamoylated lysine • Mg++brings together the reactants in a correct orientation, and stabilizes the negative charge that forms upon the nucleophilic attack of enediolate to CO2
Rubisco is Activated via Covalent Modification of the Active Site Lysine
Synthesis of Glyceraldehyde-3 Phosphate (First Stage) Three rounds of the Calvin cycle fix three CO2 molecules and produce one molecule of 3-phosphoglycerate
Fate of Glyceraldehyde 3-phosphate (Second Stage) • Converted to starch in the chloroplast • Converted to sucrose for export • Recycled to ribulose 1,5-bisphosphate