Download
1 / 33
340 likes | 627 Views
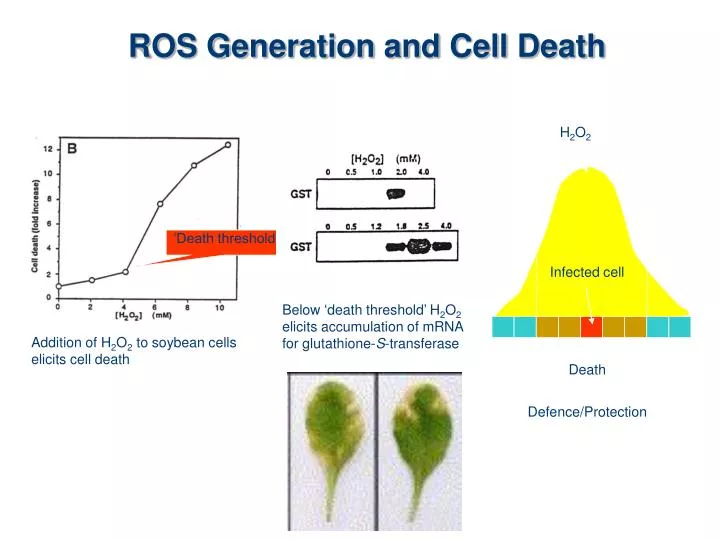
H 2 O 2. Infected cell. Death. Defence/Protection. ROS Generation and Cell Death. ‘Death threshold’. Below ‘death threshold’ H 2 O 2 elicits accumulation of mRNA for glutathione- S -transferase. Addition of H 2 O 2 to soybean cells elicits cell death. ROS are Essential.

E N D
H2O2 Infected cell Death Defence/Protection ROS Generation and Cell Death ‘Death threshold’ Below ‘death threshold’ H2O2 elicits accumulation of mRNA for glutathione-S-transferase Addition of H2O2 to soybean cells elicits cell death
ROS are Essential • thyroid cells must make H2O2 to attach iodine for Thyroxine synthesis • In plants, ROS are needed for root growth and stomata regulation • Macrophages and plants generate ROS to kill bacteria in phagosomes • NADPH Oxidase catalyzes the synthesis of the superoxide anion. NADPH − 2 e- + 2O2 -> NADP+ + H+ + 2 .O2− • This produces a large increase in oxygen consumption, called the "respiratory burst". • SOD converts it into H2O2, which kills bacteria (except which make enoughcatalase). • the enzyme myeloperoxidase catalyzes the reaction of H2O2 with Cl- to produce the strongly antiseptic hypochlorite ion (OCL−, #6 above).H2O2+ Cl− -> HOCL (hypochlorous acid) + OH−HOCL -> H+ + OCl−
rhd2 a, WT (bar, 100 µm). b, rhd2 mutant (bar, 50 µm). c, RHD2 is located on chromosome 5 d, section, showing the epidermis (EP) (bar, 25 µm). e, In situ hybridization of RHD2/AtrbohC sense probe (control). f, Antisense RHD2/AtrbohC probe. g, In situ hybridization of RHD2/AtrbohC sense (control). h,Antisense RHD2/AtrbohC probe in the region where root hairs grow i, In situ hybridization sense (control) j, Antisense RHD2/AtrbohC probe shows that RHD2 is expressed in the elongation zone.
rhd2 ROS accumulation (CM-H2DCF imaging) during root-hair elongation: transmission (top) and pseudocolour fluorescent images (bottom) are displayed for WT (a), rhd2 (b) and WT (c) after treatment with DPI
The regulation of stomatal closure ROS are essential signals mediating stomatal closure induced by ABA via the activation of calcium-permeable channels in the plasma membrane [2. ;12.]. The phenotypes of both the open stomata1 (ost1) protein kinase mutant, which is disrupted in ABA-induced ROS production but is still able to close stomata in response to H2O2, and the ABA-insensitive1-1 (abi1-1) mutant, which cannot use ABA to activate the OST1 kinase, suggest that protein phosphorylation by OST1 functions between ABI1 and ROS production in the ABA signalling cascade [13. and 14.]. Whether OST1 regulates ROS production directly via the NADPH oxidase remains unknown. Phosphatidylinositol 3-phosphate (PI3P), which has been implied to regulate stomata closure [14.], may also act via an ABA-induced generation of ROS [15.]. A.M. Hetherington and F.I. Woodward, The role of stomata in sensing and driving environmental change. Nature424 (2003), pp. 901–908. Abstract-BIOSIS Previews |12. Z.M. Pei, Y. Murata, G. Benning, S. Thomine, B. Klusener, G.J. Allen, E. Grill and J.I. Schroeder, Calcium channels activated by hydrogen peroxide mediate abscisic acid signalling. Nature406 (2000), pp. 731–734. Abstract-MEDLINE | Cell expansion and development Recent work has revealed that ROS production by plasma-membrane NADPH oxidases seems not to be limited to the control of stomata opening and defence responses. The generation of ROS may be of more general importance during plant signalling and development. For instance, the inhibition of root elongation by ABA is reduced in atrbohD atrbohF double mutants and in atrbohF mutants [4.••]. Furthermore, the atrbohD atrbohF double mutant also shows reduced inhibition of seed germination by ABA and a reduction in the induction of ROS production by bacterial The sensing of ROS ROS produced in different subcellular compartments influence the expression of a large number of genes [2.]. This suggests that cells have evolved strategies to utilise ROS as biological signals that control various genetic stress programs. This interpretation is based on the unstated assumption that a given ROS can interact selectively with a target molecule that perceives the increase of ROS concentration, and then translates this information into a change of gene expression. Such a change in transcriptional activity may be achieved through the oxidation of components of signalling pathways that subsequently activate transcription factors (TFs) or by modifying a redox-sensitive TF directly. H2O2 activates Arabidopsis thaliana MAPK3 (AtMPK3) and AtMPK6 via the activity of the MAPKKK Arabidopsis NPK1-RELATED PROTEIN KINASE1 (ANP1) [20.] and strongly induces Arabidopsis thaliana NUCLEOTIDE DIPHOSPHATE KINASE2 (AtNDPK2) [21.]. Plants that overexpress AtNDPK2 have reduced levels of ROS and enhanced tolerance of cold, salt and ROS stress, whereas the atndpk2 knockout mutant is more sensitive to oxidative stress. AtNDPK2 overexpression leads also to an increased expression of antioxidant genes
Generation of ROS during gravitropism A, Time course of gravistimulation-induced ROS generation. The roots were oriented horizontally for the indicated time and then cut into two parts. Zone 1 (black bars) contains the apical end of root to 0.4 cm, and zone 2 (hatched bars) contains 0.4 to 0.8 cm from root tip. SA, Salicylic acid-treated maize roots. Plant Physiol. 126, 1055-1060.
Auxin-induced ROS generation A, Agar blocks were incubated in IAA and then placed on the indicated region. B, Transient generation of ROS by auxin. Intracellular ROS generation in protoplast by flow cytometry. Shaded area is control fluorescence. Protoplasts incubated with IAA for 5 min (thin line), 10 min (thick line), 20 min (dashed line), or 30 min (dot line). C, Effect of NPA on ROS-induced gravitropism. Effect of asymmetric application of H2O2 on root curvature pretreatment with NAC suppressed the gravitropic curvature
OXI1 kinase activity is itself also induced by H2O2in vivo. Figure 1 H2O2, cellulase and wounding increase the expression of OXI1. a, Induction of OXI1 in seedlings treated with H2O2 (10 mM) or water. b, Induction of OXI1 by cellulase (0.01% w/v) or wounding. OXI1 expression levels were determined by northern analysis. c, Seedlings containing an OXI1::GUS gene construct were incubated in water (left panel) or 0.1% w/v cellulase solution. In the right panel, cotyledons were cut across the surface with a sharp razor blade (middle panel). After 3 h, tissues were stained for GUS expression Transient vs. prolonged Plants containing an OXI1::GUS gene construct were wounded with tweezers and stained for (a) H2O2 production with diaminobenzidine (DAB) or (b) expression of the GUS protein24 Supplement Fig.1
the specific function of OXI1 in ROS-mediated responsesOXI1 is necessary for basal resistance to P. parasitica. Figure 2a, Plants containing an OXI1::GUS gene construct were infected with virulent P. parasitica and stained for GUS expression 7 days after infection. Infected leaves show induction along fungal hyphae (top panel) and in cells containing fungal haustoria (middle panel, close-up in bottom panel). b, Infection levels of oxi1 and Ws-2 wild-type seedlings 7 days after infection. c, Seedlings were infected with Emco5 and the level of fungal infection was assessed by determining the extent of sporulation on wild-type, oxi1 mutant and oxi1 complemented with wild-type OXI1 (oxi1 + OXI1) 6 days after infection. consistent with AOS production during pathogenesis necessity of OXI1 for resistance to a virulent pathogen Resistance to the avirulent isolate Emoy2 was not removed in the oxi1 mutant (data not shown). Because AOS are also produced in response to avirulent fungi, this implies that OXI1 either is not involved in host-specific resistance or is a redundant component in this context. mutated reconstit
OXI1 is necessary for normal root hair development a, Seedlings containing an OXI1::GUS gene construct were grown on 0.8% agar and subsequently stained for GUS expression. GUS was always expressed in the root, especially high in root hairs b, Visual comparison of oxi1 and Ws-2 wild-type roots grown through air. c, Distribution of mature root hair lengths of oxi1 seedlings (filled bars) and Ws-2 wild-type seedlings (open bars) with roots growing through air. For complementation data, see Supp Fig. 3. Cessation of root hair growth is a characteristic of some rhd2 mutants14 that fail to generate necessary AOS during root hair development7 AOS are produced in response to stress and during root hair development, pointing to the OXI1 role in responses involving AOS. consistent with the necessity of an AOS signal for normal root hair cell developmentForeman, J. et al. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 422, 442–446 (2003)7
The Approach • Differential display of kinases that are induced (i.e. transcriptional control) by oxidative stress • by using degenerate primers complementary to sequences conserved between protein kinases: one gene was named OXI1, for OXIDATIVE SIGNAL-INDUCIBLE1.
ROS signal transduction pathways ROS influence many genesby translating ROS concentration into a change in transcriptional activity through the oxidation of signaling pathway components that subsequently activate transcription factors (TFs) or by directly modifying redox-sensitive TFs Mitochondria The 1st line of ROS response: ROS perception
OXI1 kinase activity in vivoOXI1 kinase activity is induced by H2O2 and cellulase,OXI1 is necessary for MPK3 and MPK6 activation. min OXI1 is activated in response to H2O2 a, Cellulase and H2O2 induce activity of the native OXI1 kinase, immunoprecipitated with an anti-OXI1 antibody from Arabidopsis leaves. b, c, MPK3 (b) and MPK6 (c) activity in cell extracts, assessed by immunocomplex kinase assays. Samples were treated with H2O2 (10 mM) or cellulase (0.1% w/v). Myelin basic protein was used as a substrate. A, activity; P, protein levels, determined by western blotting with antibodies specific for OXI1 (a), MPK3 (b) and MPK6 (c). OXI1 activates downstream MAPKS
further conclusions Downstream (indirect) targets of OXI1 are likely to include MPK3 and MPK6,because OXI1 is required for full activation of these kinases. OXI1 gene expression is also induced by cold,by osmotic stress and by heat (data not shown). It is therefore possible that OXI1 is involved in many more AOS-dependent processes and might act as a universal mediator of oxidative bursts and stresses in Arabidopsis. However,….
Functional analysis of oxidative stress-activated mitogen-activated protein kinase cascade in plants Yelena Kovtun,Wan-Ling Chiu,Guillaume Tena,and Jen Sheen* PNAS (received for review December 1,1999) • Approach: • To determine whether H2O2 signaling is mediated through an evolutionarily conserved MAPK cascade, we performed a MAPK activity in-gel assay • To determine the molecular identity and define the role of H2O2-activated MAPKs,we initiated a search for candidates that might participate in the oxidative stress-induced MAPK cascade.We chose to analyze MAPKKKs because they are the first conserved enzymes in the MAPK cascade • To elucidate the function of ANPs,we first demonstrated that ectopically expressed ANPs could activate endogenous MAPKs, ANP1 initiates an H2O2-activated MAPK cascade. • ANP1 Activates H2O2-Inducible Promoters. To provide further evidence for the specific involvement of ANPs in H2O2 signaling and to investigate their downstream targets, we tested the effect of constitutively active ANP1 on the activity of the GST6 • Constitutively Active NPK1 enhances tolerance to multiple stresses in transgenic tobacco. Oxidative stress-activated GSTs and HSPs encode conjugation enzymes and molecular chaperones, respectively. They play essential roles in detoxification and stabilization
Flooding causes anoxia and an anaerobiotic response in roots.
Oxygen sensing and plant responses to hypoxia Aerenchyma formation in maize roots during flooding Calcium fluctuations in maize roots during hypoxia
Sensing oxygen and its derivatives • In bacteria O2.-and H2O2 induce specific operons that contain antioxidant genes via direct oxidation of the transcription factor • oxyR regulates a group of enzymes that defend against oxidative stress. Constitutive oxyR mutants overproduce 9 proteins that are induced by oxidative stress, including MnSOD, and glutathione reductase • In yeast H2O2 sensing through oxidation of the Yap1 transcription factor via Cys oxidation • In animals a family of transcription factors are induced by Hypoxia (HIF) via an O2– dependend hydroxylation reaction
Oxidation/reduction states are sensed in bacteria Hybrid Kinase activates the ArcB kinase Amplification loop Transcription factor Antioxidant genes
H2O2 in plant cells H2O2, entering cells from the apoplast or generated internally, diffuses mainly within sub-cellular microdomains Depending on the H2O2 amount and the antioxidant status. H2O2 can oxidize or otherwise modulate signaling proteins, such as protein phosphatases (PP), kinases (PK) including a plasma membrane histidine kinase and MAPK cascades, TFs, and calcium channels that are located in the plasma membrane or elsewhere.
Asc, GSH, and H2O2 function as upstream/downstream components of hormone-mediated signal transduction &hormones may induce processes that produce H2O2 Hormone organ signal molecule function Auxin Root H2O2 Gravitropism Abscisic acid Leaves H2O2 Stomatal closure Ca2+ Leaves H2O2 Stomatal closure Abscisic acid Aleurone Antioxidants Cell survival Gibberelic acid Aleurone H2O2 Programmed cell death
ROI-mediated induction of gst1 is not dependent on ethylene, SA or Me-JA. Time-course of gst1::luc induction in etr1 mutant compared with wild type Time-course of gst1::luc induction in nahG transformants compared with wt
ROI-induced gst1 and pal1 gene expression is dependent on MAPKK activity
Two of the major signal transduction pathways in plant cells involve cytosolic calcium and protein kinases. Calcium-dependent protein kinases, which are very prevalent in plant cells, connect the two transduction pathways (dashed arrow).
Plants’ way to overcome environmental stress • Genes, genes, genesSome plants know what to do different genes or different expression pattern • The proof is in the puddingResistant & susceptible speciesto activate the right genes at the right time --> Signal Transduction END
Plants have to exploit their immediate environment to maximum effect. Their inability to move means that the best way of dealing with stresses is through physiological or morphological changes. Abiotic stresses, and ways to adapt to them are numerous and interlinked.
OXI1 kinase is necessary for oxidative burst-mediated signalling in Arabidopsis ROS generated in response to stimuli and during development can function as signalling molecules, leading to specific downstream responses In plants these include such diverse processes as • coping with stress (for example: ) • pathogen attack, • Wounding, • Drought, salt, cold • oxygen deprivation (hypoxia), • ABA-induced guard-cell closure, cellular development (for example root hair growth7) Nature427, 858 - 861 (26 February 2004)
The Plant JournalVolume 37Page 877 - March 2004 overlapping gene pools biased towards 'abiotic' and 'biotic' gene pools -primarily abiotic -primarily biotic MDA originates from saturated FAs MVK is similar to MDA Conclusion: small carbonyl compounds have different activities and thus can convey regulatory information. Different ROS induce specific genes specificity in ROS recognition/signaling
Different environmental signals can affect the flow of the signalling pathway but still result in the same outcome OR in a different outcome
Two stress responses to singlet oxygen:(i) growth inhibition and (ii) lethality Comparison of singlet oxygen production, cell death, and growth inhibition in executer1/flu, flu, and WT. Etiolated seedlings of flu (B) and executer1/flu (A) overaccumulated similar amounts of Pchlide, as indicated by the bright red fluorescence, in contrast to etiolated WT (C). Cell death of flu seedlings (B) and growth inhibition of mature flu plants (=) grown under nonpermissive 16 h light–8 h dark conditions were blocked by the executer1 mutation (■) (D and E). Results from WT control plants (▲) are shown in (D) and (E). (F to H) Generation of singlet oxygen. WT (F), flu (G), and executer1/flu (H) were grown under continuous light until bolting. Then plants were either transferred to the dark for 8 h or kept under light. Cut leaves were infiltrated with dansyl-2,2,5,5-tetramethyl-2,5-dehydro-1H-pyrrole and subsequently illuminated with white light. Singlet oxygen trapping was measured as relative quenching of DanePy fluorescence. In WT controls, no difference in singlet oxygen production between continuous light and a dark-light shift was found (H), whereas in leaves of flu and executer1/flu generation of singlet oxygen was enhanced in plants that had been kept in the dark before reillumination ( ) but not in plants exposed to continuous light ( ). ex1, executer1.