Download
1 / 70
760 likes | 1.23k Views
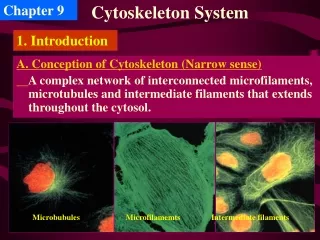
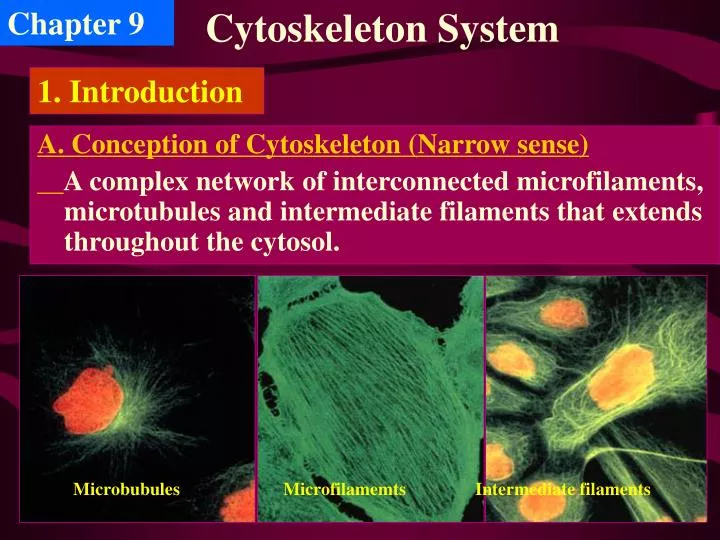
Chapter 9. Cytoskeleton System. 1. Introduction. A. Conception of Cytoskeleton (Narrow sense) A complex network of interconnected microfilaments, microtubules and intermediate filaments that extends throughout the cytosol. Microbubules. Microfilamemts. Intermediate filaments.

E N D
Chapter 9 Cytoskeleton System 1. Introduction A. Conception of Cytoskeleton (Narrow sense) A complex network of interconnected microfilaments, microtubules and intermediate filaments that extends throughout the cytosol. Microbubules Microfilamemts Intermediate filaments
Fluorescent microscopy and Electron microscopy : Immunofluorescence: fluorescently-labeled antibody Fluorescence: microinject into living cells Video microscopy: in vitro motility assays Electron: Triton X-100, Metal replica Quick freezing-deep etching EM • Biochemical analysis (in vitro) Difference centrifugation; SDS-PAGE • Drugs and mutations (about functions) B. Techniques for studying the cytoskeleton
C. The self-assembly and dynamic structure of cytoskeletal filaments • Each type of cytoskeletal filament is constructed from smaller protein subunits. • The cytoskeleton is a network of three filamentous structures. • The cytoskeleton is a dynamic strucrure with many roles.
2. Microfilament, MF • Using ATP, G-actin polymerizes to form MF(F-actin) A. MFs are made of actin and involved in cell motility.
Characteristics: (1) Within a MF, all the actin monomers are oriented in the same direction, so MF has a polarity B. MF assembly and disassembly Myosin is molecular motor for actins.
(2) In vitro, (Polymerization) both ends of the MF grow, but the plus end faster than the minus. Because actin monomers tend to add to a filament ’ s plus end and leave from its minus end----
(3) Dynamic equilibrium between the G-actin and polymeric forms, which is regulated by ATP hydrolysis and G-actin concentration.
2.2 Assembly ◆ Mechanism of actin polymerization: 3 phases of G-actin polymerization. ◆ Critical concentration (Cc). In steady state, G-actin monomers only exchange with subunits at the filament ends but there is no net change in the total mass of filaments. ◆ During the elongation state, one end of the filament, the (+) end, elongates five to ten times faster than does the opposite (-) end. This is because Cc value is much lower for G-actin addition at the (+) end than for addition at the (-) end.
Figure 6-17 The three phases of G-actin polymerization in vitro.
(4) Dynamic equilibrium is required for the cell functions. Some MFs are temporary and others permanent.
(5)The nucleation of actin filaments at the PM is frequently regulated by external signals, allowing the cell to change its shape and stiffness rapidly in response to changes in its external environment. This nucleation is catalyzed by a complex of proteins that includes two actin-related proteins, or ARPs(Arp2 and Arp3).
Cytochalasins: Prevent the addition of new monomers to existing MFs, which eventually depolymerize. C. Specific drugs affect polymer dynamics Phalloidin: A cyclic peptide from the death cap fungus, blocks the depolymerization of MF Those drugs disrupt the monomer-polymer equilibrium, so are poisonous to cells
The structures and functions of cytoskeleton are mainly controlled by its binding proteins D. Actin-binding proteins
2.4 microfilament-binding proteins Actin binding proteins control the structure and behavior of actin filament. ◆ actin binding proteins e.g. proflin (promote acting assembly), thymosin beta4 (inhibits actin assembly). Some cytosolic proteins control actin polymerization. ◆ microfilament-binding proteins ◆ 3 different types of stalbe actin filament structures: ◆Parallel bundle: MFs isotactic parallel arrange,mainly found in microvillus and filopodium (丝状伪足). ◆Contractile bundle: MFs anti-parallel arrange, mainly found in stress fibers (应力纤维) and contractive ring of mitosis(有丝分裂收缩环 )。 ◆Gel-like network: MFs cross-linked arrange, most be found in cell cortex (cytosol, 细胞皮层).
成核蛋白(nucleating proteins): actin-related proteins, ARPs 单体–隔离蛋白(monomer-sequestering protein):thymosin 封端(加帽)蛋白(End-blocking(capping) proteins):capZ 单体–聚合蛋白(monomer-polymerizing proteins):抑制蛋白(profilin)是一种与ATP-肌动蛋白单体结合的蛋白质 肌动蛋白纤维解聚蛋白(actin filament-depolymerizing proteins): cofilin、ADF以及蚕食蛋白与肌动蛋白纤维的减端结合,大大促进肌动蛋白纤维解聚成单体。 交联蛋白(cross-linking proteins): ABP280和细丝蛋白,促进形成近于正交相互联系的纤维松散网络 纤维–切割蛋白(filament-severing proteins): gelsolin 膜结合蛋白(membrane-binding proteins):连接膜与肌动蛋白的蛋白质包括联结蛋白(vinculin),ERM家族的成员包括埃兹蛋白(ezrin)、根蛋白(radixin)和膜突蛋白(moesin)
Model of the complementary roles of profilin and thymosin β4 in regulating polymerization of G-actin.
Actin filaments are likewise strongly affected by the binding of accessory proteins along their sides. • Actin filaments in most cells are stabilized by the binding of tropomyosin, an elongated protein. Which can prevent the filament from interacting with other proteins. • Another important actin filament binding protein, cofilin, present in all eucaryotic cells, which destabilized actin filaments(also called actin depolymerizing factor). • Cofilin binds along the length of the actin filament, forcing the filament to twist a little more tightly. • In addition, cofilin binding cause a large increase in the rate of actin filament treadmilling.
The modular structures of four actin-cross-linking proteins • The formation of two types of actin filament bundles: • Contractile bundle mediated by α-actinin • parallel bundle mediated by fimbrin.
Actin filaments are often nucleated at the plasma membrane. The highest density of actin filaments is at the cell periphery forming cell cortex. • Gel-like network Filamin cross-links actin filaments into a three-dimensional network with the physical properties of a gel. Loss of filamin causes abnormal cell motility
E. Functions of MFs (1) Maintain cell ’ s shape and enforce PM
Platelet activation is a controlled sequence of actin filament severing,uncapping, elongation,recapping, and cross-linking. (2) Cell migration (Fibroblast et al)
(3) Microvillus: Support the projecting membrane of intestinal epithelial cells
Composed of actin filaments and myosin-II (4) Stress fibers Focal contacts MFs Focal contacts Stress Fibers Response to tension Response to tension
Organization of skeletal muscle tissue (6) Muscle contraction
皮肤 ?蛋白 平原皮肤球 ??蛋白 同伴皮肤 ?蛋白
Myosin: The actin motor portein • Proteins play important roles in muscle contraction Myosin II--Dimer ATPase Mainly in muscle cells Thick filamemts
Light-chain phosphorylation and the regulation of the assembly of myosin II into thick filaments
Tropomyosin, Tm and Tropnin, Tn Ropelike molecule Complex, Ca2+-subunit Regulate MF to bind to the head of myosin Control the position of Tm on the surface of MF
Action potential • Excitation-contraction • coupling process Ca2+ rise in cytosol Tn Tm Sliding
Schematic diagram showing how a Ca2+-release channel in the sarcoplasmic reticulum membrane is thought to be opened by a voltage-sensitive transmembrane protein in the adjacent T-tubule membrane
Smooth and nonmuscle cell contraction are regulated in a manner distinct from that of skeletal muscle cells F. Smooth muscle cell(平滑肌细胞 ) contraction Bind to MLCK Ca2+ rise Ca2+ -calmodulin Regulate light chain Phosphorylate Myosin interact with actin Contraction SLOW
3.Microtubule, MT A. Structures: Tubulin heterodimers are the protein building blocks of MTs
Singlet Double Triplet Arrangement of protofilaments in singlet, double, and triplet MTs A A B B C In cilia and flagella In centrioles and basal bodies
(1) Interphase: Centrosome B. MTs assemble from microtubule-organizing centers (MTOCs) Dynamic instability (2) Dividing cell: Mitotic spindle Dynamic instability (3) Ciliated cell: Basal body Stability
Dynamic instability due to the structural differences between a growing and a shrinking microtubule end. • GTP cap; • Catastrophe: accidental loss of GTP cap; • Rescue: regain of GTP cap C. Characteristics of MT assembly
Structure • Why the centrosome can act as MTOC No centrioles in Plant and fungi
MT are nucleated by a protein complex containing ?-tubulin The centrosome is the major MTOC of animal cells
(1) Colchicine • Drugs affect the assembly of MTs Binding to tubulin dimers, prevent MTs polymerization (2) Taxol Binding to MTs, stabilize MTs These compounds are called antimitotic drugs, and have application in medical practice as anticancer drugs
MAPs modulate MT structure, assembly, and function • Microtuble-associated proteins (MAPs) Control organization Motor MAPs Nonmotor MAPs Tau: In axon, cause MTs to form tight bundles MAP2: In dendrites, cause MTs to form looser bundles
The importance of MAPs for neurite formation Like dendrite Like axon
Organization of MT bundles by MAPs . Spacing of MTs depends on MAPs Insect cell expressing MAP2 Insect cell expressing tau From J. Chen et al. 1992. Nature 360: 674
The effects of proteins that bind to MT ends (A)The transition between Mt growth and Mt shrinking is controlled in cells by special proteins. ??蛋白 (B)Capping proteins help to localize Mt in budding yeast cell.
5. Functions of MTs 1. Maintain cell shape