Download
1 / 1
10 likes | 100 Views
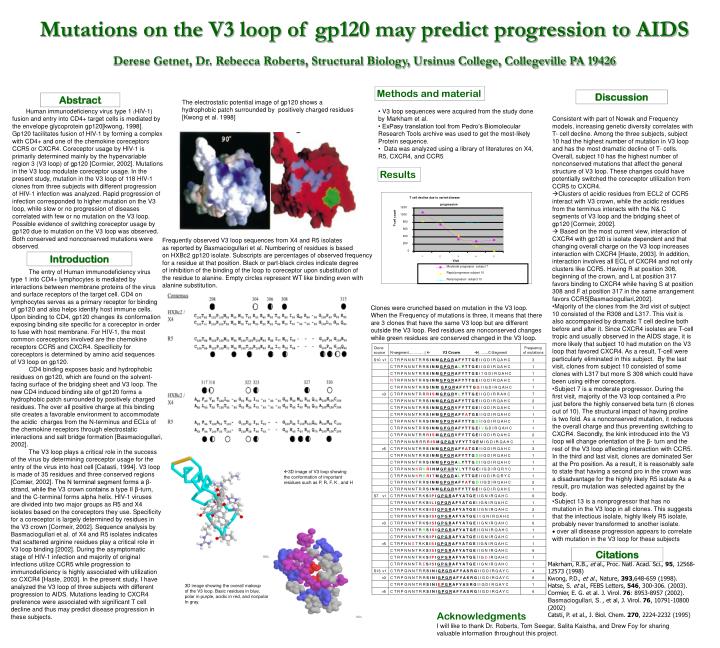
T cell decline due to varied disease. progression. 1200. 1000. T-cell count. 800. 600. 400. 200. 0. 1. 2. 3. 4. 5. Visit. Moderate progressor subject 7.

E N D
T cell decline due to varied disease progression 1200 1000 T-cell count 800 600 400 200 0 1 2 3 4 5 Visit Moderate progressor subject 7 The electrostatic potential image of gp120 shows a hydrophobic patch surrounded by positively charged residues [Kwong et al. 1998] Rapid progressor subject 10 Nonprogressor subject 13 Frequently observed V3 loop sequences from X4 and R5 isolates as reported by Basmaciogullari et al. Numbering of residues is based on HXBc2 gp120 isolate. Subscripts are percentages of observed frequency for a residue at that position. Black or part-black circles indicate degree of inhibition of the binding of the loop to coreceptor upon substitution of the residue to alanine. Empty circles represent WT like binding even with alanine substitution. Mutations on the V3 loop of gp120 may predict progression to AIDS Derese Getnet, Dr. Rebecca Roberts, Structural Biology, Ursinus College, Collegeville PA 19426 Methods and material Discussion Abstract Human immunodeficiency virus type 1 (HIV-1) fusion and entry into CD4+ target cells is mediated by the envelope glycoprotein gp120[kwong, 1998]. Gp120 facilitates fusion of HIV-1 by forming a complex with CD4+ and one of the chemokine coreceptors CCR5 or CXCR4. Coreceptor usage by HIV-1 is primarily determined mainly by the hypervariable region 3 (V3 loop) of gp120 [Cormier, 2002]. Mutations in the V3 loop modulate coreceptor usage. In the present study, mutation in the V3 loop of 118 HIV-1 clones from three subjects with different progression of HIV-1 infection was analyzed. Rapid progression of infection corresponded to higher mutation on the V3 loop, while slow or no progression of diseases correlated with few or no mutation on the V3 loop. Possible evidence of switching coreceptor usage by gp120 due to mutation on the V3 loop was observed. Both conserved and nonconserved mutations were observed. • V3 loop sequences were acquired from the study done • by Markham et al. • ExPasy translation tool from Pedro’s Biomolecular • Research Tools archive was used to get the most-likely • Protein sequence. • Data was analyzed using a library of literatures on X4, • R5, CXCR4, and CCR5 • Consistent with part of Nowak and Frequency models, increasing genetic diversity correlates with T- cell decline. Among the three subjects, subject 10 had the highest number of mutation in V3 loop and has the most dramatic decline of T- cells. Overall, subject 10 has the highest number of nonconserved mutations that affect the general structure of V3 loop. These changes could have potentially switched the coreceptor utilization from CCR5 to CXCR4. • Clusters of acidic residues from ECL2 of CCR5 interact with V3 crown, while the acidic residues from the terminus interacts with the N& C segments of V3 loop and the bridging sheet of gp120 [Cormeir, 2002]. • Based on the most current view, interaction of CXCR4 with gp120 is isolate dependent and that changing overall charge on the V3 loop increases interaction with CXCR4 [Haste, 2003]. In addition, interaction involves all ECL of CXCR4 and not only clusters like CCR5. Having R at position 308, beginning of the crown, and L at position 317 favors binding to CXCR4 while having S at position 308 and F at position 317 in the same arrangement favors CCR5[Basmaciogullari,2002]. • Majority of the clones from the 3rd visit of subject 10 consisted of the R308 and L317. This visit is also accompanied by dramatic T cell decline both before and after it. Since CXCR4 isolates are T-cell tropic and usually observed in the AIDS stage, it is more likely that subject 10 had mutation on the V3 loop that favored CXCR4. As a result, T-cell were particularly eliminated in this subject. By the last visit, clones from subject 10 consisted of some clones with L317 but more S 308 which could have been using either coreceptors. • Subject 7 is a moderate progressor. During the first visit, majority of the V3 loop contained a Pro just before the highly conserved beta turn (6 clones out of 10). The structural impact of having proline is two fold. As a nonconserved mutation, it reduces the overall charge and thus preventing switching to CXCR4. Secondly, the kink introduced into the V3 loop will change orientation of the β- turn and the rest of the V3 loop affecting interaction with CCR5. In the third and last visit, clones are dominated Ser at the Pro position. As a result, it is reasonably safe to state that having a second pro in the crown was a disadvantage for the highly likely R5 isolate As a result, pro mutation was selected against by the body. • Subject 13 is a nonprogressor that has no mutation in the V3 loop in all clones. This suggests that the infectious isolate, highly likely R5 isolate, probably never transformed to another isolate. • over all disease progression appears to correlate with mutation in the V3 loop for these subjects Results Introduction The entry of Human immunodeficiency virus type 1 into CD4+ lymphocytes is mediated by interactions between membrane proteins of the virus and surface receptors of the target cell. CD4 on lymphocytes serves as a primary receptor for binding of gp120 and also helps identify host immune cells. Upon binding to CD4, gp120 changes its conformation exposing binding site specific for a coreceptor in order to fuse with host membrane. For HIV-1, the most common coreceptors involved are the chemokine receptors CCR5 and CXCR4. Specificity for coreceptors is determined by amino acid sequences of V3 loop on gp120. CD4 binding exposes basic and hydrophobic residues on gp120, which are found on the solvent-facing surface of the bridging sheet and V3 loop. The new CD4 induced binding site of gp120 forms a hydrophobic patch surrounded by positively charged residues. The over all positive charge at this binding site creates a favorable environment to accommodate the acidic charges from the N-terminus and ECLs of the chemokine receptors through electrostatic interactions and salt bridge formation [Basmaciogullari, 2002]. The V3 loop plays a critical role in the success of the virus by determining coreceptor usage for the entry of the virus into host cell [Catasti, 1994]. V3 loop is made of 35 residues and three conserved regions [Comier, 2002]. The N terminal segment forms a β-strand, while the V3 crown contains a type II β-turn, and the C-terminal forms alpha helix. HIV-1 viruses are divided into two major groups as R5 and X4 isolates based on the coreceptors they use. Specificity for a coreceptor is largely determined by residues in the V3 crown [Cormeir, 2002]. Sequence analysis by Basmaciogullari et al. of X4 and R5 isolates indicates that scattered arginine residues play a critical role in V3 loop binding [2002]. During the asymptomatic stage of HIV-1 infection and majority of original infections utilize CCR5 while progression to immunodeficiency is highly associated with utilization so CXCR4 [Haste, 2003]. In the present study, I have analyzed the V3 loop of three subjects with different progression to AIDS. Mutations leading to CXCR4 preference were associated with significant T cell decline and thus may predict disease progression in these subjects. Clones were crunched based on mutation in the V3 loop. When the Frequency of mutations is three, it means that there are 3 clones that have the same V3 loop but are different outside the V3 loop. Red residues are nonconserved changes while green residues are conserved changed in the V3 loop. 3D image of V3 loop showing the conformation of important residues such as P, R, F, K , and H Citations Makrham, R.B., et al., Proc. Natl. Acad. Sci., 95, 12568-12573 (1998) Kwong, P.D., et al.,Nature, 393,648-659 (1998). Hatse, S. et al., FEBS Letters, 546, 300-306. (2003). Cormier, E. G. et al. J. Virol. 76: 8953-8957 (2002). Basmaciogullari, S., et al, J. Virol. 76, 10791-10800 (2002) Catsti, P. et al., J. Biol. Chem. 270, 2224-2232 (1995) 3D image showing the overall makeup of the V3 loop. Basic residues in blue, polar in purple, acidic in red, and nonpolar In gray. Acknowledgments I will like to thank Dr. Roberts, Tom Seegar, Salita Kaistha, and Drew Foy for sharing valuable information throughout this project.