Download

1 / 35
350 likes | 389 Views
Macroevolution or - the evolution of species. The Biological Species Concept: A species is a population or group of populations whose members have the potential to interbreed in nature to produce fertile offspring, and whose members are reproductively isolated from other

E N D
Macroevolution or - the evolution of species The Biological Species Concept: A species is a population or group of populations whose members have the potential to interbreed in nature to produce fertile offspring, and whose members are reproductively isolated from other such groups.
But this is a hellish definition to test species… Darwin (1853): “ After describing a set of forms as distinct species; tearing up my [manuscript], & making them one species; tearing that up & making them separate, & then making them one again (which has happened to me) I have gnashed my teeth, cursed species, & asked what sin I committed to be so punished.” (Darwin was trying to separate barnacles.)
More practical problems: 1) How do we deal with asexual species (bacteria, many fungi, some fish, and assorted others)? They do not interbreed. They don’t fit the definition. 2) How do we deal with fossils? Are morphological differences sufficient to define species? That would not always work with living species. 3) Does a different morphology necessarily mean a different species? No! 4) What about geographically separate populations? How can we tell if they can interbreed “in nature”? The species concept may answer this question.
What happens when apparently different species are brought together artificially (in zoos, etc.)? Lions and tigers interbreed in zoos, producing ligers. Bison and cattle breed on farms, producing commercially valuable ‘beefalos’. It only works successfully in one direction: bull with buffalo cow Why?
Note the front shoulders of this beefalo. A female cow can’t handle birthing the calf of the cross without dangerous distress.
So, how do new species evolve? Through accumulation of genetic differences. Eventually, sufficient difference has accumulated to reproductively isolate the two groups, even if they should be in contact. Those differences usually accumulate while the groups (populations) are geographically isolated.
An example from northern Canada: There are two major species of lemmings (collared lemming, brown lemming). Lemmings lived in the shadow of receding glaciers. As James Bay opened with glacial melting, the western and eastern populations of lemmings were separated (geographically isolated). Differences accumulated over time producing the two lemming species we have today. However, there are other mechanisms that can produce reproductive isolation even while groups are in close proximity. These barriers are separated into pre-zygotic (preventing zygote formation) and post-zygotic (affecting embryo development).
Pre-zygotic barriers: 1) Habitat isolation - e.g., marsh versus forest warblers, or parasites limited to different host species 2) Temporal isolation - e.g., breeding seasons in yellow-headed (May - early June) and red-winged (late June - early July) blackbirds. 3) Behavioural isolation - it’s the species specific mating dances that many of the 400 Hawaiian Drosophila do in courtship that reproductively isolates them. 4) Mechanical isolation - e.g. imagine Great Danes and chihuahuas attempting to mate
5) Gametic isolation - sperm (or pollen) and egg must be chemically compatible. In plants, pollen doesn’t germinate, pollen tubes fail to grow,… In animals, membrane proteins don’t match, and fertilization doesn’t occur
post-zygotic barriers 1) hybrid inviability - genetically programmed development is a complicated process. Species differ in this program. When genomes are mixed in a hybrid, conflicts result in the embryo failing to develop completely. They generally don’t survive. 2) hybrid sterility - the hybrids survive to maturity, but cannot produce viable offspring. The reason is usually traceable to incompatible chromosomes that don’t match up in meiosis. An example: horses and donkeys mate, the offspring (mules) are viable, but sterile
3) hybrid breakdown - first generation hybrids are viable and fertile, but the second generation (or beyond) are feeble (low survivorship and greatly reduced reproductive output) or sterile. This occurs among different species of cotton. In the absence of successful barriers to hybridization - gene exchange between ‘species’ occurs. It is called introgression. With gene flow, new species can’t form. So, think of populations in the process of accumulating differences never being able to accumulate sufficient differences to speciate.
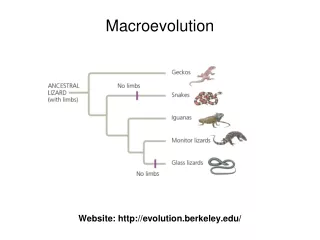
Even though various mechanisms can lead to reproductive isolation, the most common remains geographic isolation. The separation can occur at three ‘levels’: 1. Allopatry - allopatric speciation 2. Parapatry - parapatric speciation 3. Sympatry - sympatric speciation
Allopatric speciation - A population becomes geographically fragmented A body of water (river, ocean) may separate them (e.g. the lemmings). A small group may colonize an island (e.g. Darwin’s finches Plate tectonics may cause the rise of mountains between them. - Either due to environmental differences between sites (differences in regimes of natural selection) or chance events in small colonist groups (drift) genetic differences between groups accumulate.
- Eventually, sufficient differences accumulate to • prevent interbreeding. At this point we say a • new species has evolved. • - Differences appear and spread more rapidly in • small populations (drift!; mutation is not more • likely in small populations) • Frequently, it is marginal populations within • what had been a large, widespread population • that become isolated (more likely to encounter • different environments). • - Adaptive radiation may occur as small groups • become repeatedly isolated, e.g. Darwin’s • finches or Hawaiian silverswords.
A ground finch Sharp-beaked finch (Geospiza nebulosa) A cactus finch Small cactus finch (Geospiza scandens) Another ground finch with a smaller bill Small ground finch (G. fuliginosa)
A few of the wild and wonderful Hawaiian silver- swords. All grow at upper elevations in Hawaii. The most remarkable is the Haleakala silversword, which grows in the cone of an ‘active’ volcano on Maui. Haleakala silversword, Argyroxiphium sandwichense
Island adaptive radiation can go in strange directions. These are ‘sunflowers’ that result from adaptive radia- tion on the island of St.Helena in the south Atlantic.
Parapatric speciation - Populations are not separated; their boundaries contiguous - Speciation can occur when a strong environmental gradient extends across the boundary between populations - Differences in selection pressures must be great enough to overwhelm any gene flow across the boundary - Two examples: distribution over a mountainside distribution over a mine spoil gradient
Imagine a mountainside. It gets colder as you climb. Temperature changes by 10°C per kilometer. One species lives at the bottom of the slope, the other at the top. Differences in temperature adaptations may mean that reproductive success is very much lower in the ‘wrong’ part of the slope. Slowly, populations come to differ in many ways, and parapatric speciation occurs.
The other example is of grasses growing on and off land of a mine in Wales that produced lead. Heavy metals (lead, copper, nickel) are poisonous to many (most) plants. Selection on mine lands produced a variety of Agrostis tenuis that was tolerant of lead. A part of the change (mutation) giving tolerance was a shift in flowering time. Thus, although tolerant and intolerant plants grow in adjacent areas, there is little or no gene flow between populations, and speciation can occur.
Sympatric speciation - Populations overlap in distribution (sym - same; patra - country). Then how can they become reproductively isolated? - Two accepted ways: by host specialization by becoming polyploid - Host specialization - when host is both feeding and mating site, a change in host can isolate the shifted population. Example in text: Rhagoletis pomonella normally feeds and mates on hawthorn fruits. Some switched in NY in 1864 to feeding on apples. In 1960 some switched again, to cherries.
- becoming polyploid: autopolyploids - double chromosome number by non-disjunction or nuclear fusion in meiosis. Diploid gametes self-fertilize. Result is tetraploid. It cannot backcross with parents, but is fertile with a like type.
Allopolyploidy - fertilization involving gametes from two different species. Interspecific hybrids are usually inviable or sterile (due to failure of chromosome pairing in synapsis of prophase in meiosis; chromosomes aren’t really homologues), but… Non-disjunction in the first generation of the hybrid can make a viable, fertile hybrid.
Is allopolyploidy important? Did anyone have bread (pizza, big Mac, sandwich) for lunch? The bread was made from wheat. Allopolyploidy produced Triticum aestivum, or bread wheat. Its chromosome complement is 2n=42. That complement arose by spontaneous hybridization of 2 other wheat grasses with 28 and 14 chromosomes. 2n=28 --> n=14 in gametes 2n=14 --> n=7 seems incompatible, but by non-disjunction in meiosis of this hybrid, a fertile species with 2n=42 was formed.
Adaptive radiation occurs on continents, as well, but continental drift and plate tectonics brings faunas into contact, and there are then extinctions. Here are drawings of South American mammals driven extinct after the rise of Central America permitted exchange. Only the armadillo and opossum successfully moved north. A host of larger North American mammals crossed southward, and drove ecologically similar species extinct. The text has a diagram of the North American mammals that were (at least temporarily) successful in the south.
Here are diagrams of what they drove extinct: Probably related to condylarths, with camel- like habits Large, mastodon-like
Extinction of species is as important to what we observe today as is speciation. There is a nominal background rate of species disappearance (extinction). However, in the history of life on earth there have been periods when much larger numbers of extinctions occurred. These are called mass extinctions. There have been 5 mass extinctions (and humans are almost certainly driving a 6th. Causes of some are not known. One seems relatively well explained - the mass extinction that occurred 65 MYBP, and eliminated dinosaurs, making room for diversification and enlargement of the mammals.
The Cretaceous mass extinction was caused by a combination of climate change and the collision of a ~17km diameter asteroid into the Yucatan peninsula. The effects of collision were much like the nuclear winter that would be caused by global nuclear warfare: Intense fires over much of Mexico and the U.S., a global dust cloud darkening the skies for months, a sudden change in climate as a result, death of plants, and therefore a lack of food to support the huge vegetarian dinosaurs, leading to the death of the meat eaters.
We are driving another mass extinction. Many species are going extinct each day, though we don’t even know their names, and may not have even discovered them yet. How? We are cutting down tropical forests in South America, Africa, and Asia for lumber, firewood, and conversion to agricultural land, both on the large scale and as a result of population increase and ancient practice of slash and burn farming. The effect of this is not only loss of plant species, but loss of the diversity of insect and other species directly or indirectly dependent on the plants.
Tempo and mode in Evolution There are two views of the rate of apparent change in species: 1) the microevolutionary view - new species formation results from gradual accumulation of phenotypic (usually seen as morphological) change. 2) the punctuated equilibrium - most of the morpho- logical change becomes apparent when species initially form. Populations are then very small. Selection can rapidly move the characteristics of the entire species, and drift can lead to rapid change. Through the remainder of the species’ history, there is little evident change.
The result is a history in the fossil record of long stasis (equilibrium, constancy) punctuated by short periods of dramatic change. Is only one of these hypotheses correct, and the other wrong? No! A punctuated equilibrium is evident in the fossil record. The ‘sudden’ change may represent 1000s of years of gradual change, but it looks rapid when viewed on a geological time scale.