Download

1 / 27
320 likes | 744 Views
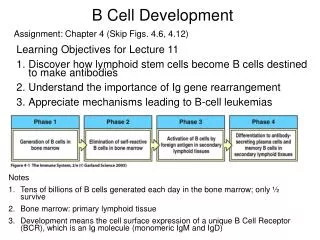
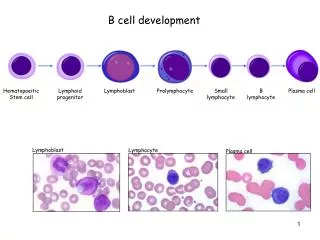
A Few More Things About B Cell Development. Cellular and Molecular Immunology Chapters 7 & 9. Allelic exclusion. Isotypic exclusion. The steps in immunoglobulin gene rearrangement at which cells can be lost. Clonal deletion-Antigen-induced loss from the B cell population

E N D
A Few More Things About B Cell Development Cellular and Molecular Immunology Chapters 7 & 9
Allelic exclusion Isotypic exclusion The steps in immunoglobulin gene rearrangement at which cells can be lost.
Clonal deletion-Antigen-induced loss from the B cell population Anergy-State of non-reactivity Binding to self molecules in the bone marrow can lead to the death or inactivation of immature B cells.
The interaction of B cells with T cells induces three important events in B cells Isotype switching Somatic hypermutation Affinity maturation
Plasma cells secrete antibody at a high rate but can no longer respond to antigen or helper T cells.
The Major Histocompatibility Complex Cellular and Molecular Immunology Chapter 4
MHC - The Major Histocompatibility Complex Originally identified in mice as blood cell antigens by R.A. Gorer and G.D. Snell in 1930s, and defined on the basis of tissue graft rejection - 1980 Nobel prize awarded to Snell. Work by Rolf Zinkernagel and Peter Doherty in the 1970s revealed that it is the complex of MHC molecule plus antigen that is recognized by T cells (MHC restriction of T cell responses) - 1996 Nobel Prize Two classes of MHC molecule: Class I (single MHC a chain + b2-microglobulin) and Class II (a chain plus b chain) - more Ig-superfamily members (Ig-C domains)
Peptide Binding to MHC Molecules: Class I vs. II Class IClass II • Generated by proteasome (cytosol) • Generated in endosomes/lysosomes • Transported to ER by TAP1/2 • Bind in specialized vessicles (MIIC) • Restricted length, 8-9 residues • Any length, extend from MHC • Binding energy from N, C termini • Binding energy from backbone • “Anchor” residues fill specific pockets • “Anchor” residues fill specific pockets
Fig. 3.27 The T-cell receptor binding to the MHC:peptide complex (Ian Wilson).
The Major Histocompatibility Complex MHC Class I and Class II genes are encoded in a large (>4 Mb, >200 genes) tightly linked gene cluster: the MHC The MHC is similarly organized in mice (Ch17) and humans (Ch6) - syntenic Known in humans as HLA (human leukocyte antigen) genes; in mouse as H-2 (histocompatibility group 2) genes Genes are highly polymorphic (many alleles) Class I: HLA-A, -B, -C (human); H-2K, -2D, -2L (mouse) Class II: HLA-DP, -DQ, -DR (human), I-A, I-E (mouse)
Human MHC class I: HLA-A, -B, -C MHC class II: HLA-DR, -DP, -DQ Mouse MHC class I: H2-K, -D, -L MHC class II H2-A, -E (I-A, I-E) b2-microglobulin is located on a different chromosome.
MHC Encodes a Variety of Other Genes Class I region: “Non-classical” Class I molecules (share sequence homology, bind b2m, but non-polymorphic), also called Class IB genes H2-M3 (mouse) binds N-formylated peptides MIC genes interact with activating receptors on NK (and some T) cells HLA-E and HLA-G interact with inhibitory receptors on NK cells HFE is involved in iron metabolism
MHC Encodes a Variety of Other Genes Class II region: other genes involved in antigen processing TAP1 and TAP2 - transporter for Class I peptides Tapasin - involved in loading peptides into Class I molecules LMP2 and LMP7 - inducible proteasome subunits HLA-DM (H-2M in mouse) and HLA-DO (H-2O) - control loading of peptides into Class II molecules (DM+, DO-) Gene expression (except DOb) is coordinately regulated by IFN-g induction of Class II Transactivator (CIITA)
MHC Encodes a Variety of Other Genes Class III region: miscellaneous assortment of genes - some immunologically related, others not Complement C4, C2, and factor B Tumor necrosis factor (TNF) and lymphotoxin (LT) 21-hydroxylase (enzyme involved in steroid biosynthesis)
MHC Polymorphism MHC Class I and Class II molecules have many alleles Diversity ensures that a wide range of peptides can be presented within the population (even if a much more limited set is presented by any individual) For Class II, both a and chains are polymorphic (except DRa in humans and Ea in mice), adding more diversity
Fig. 5.16 Allelic variation occurs at specific sites within the MHC molecules. The polymorphism of MHC guarantees a sufficient number of different MHC molecules in a single individual to present a variety of peptides.
MHC Polymorphism MHC Class I and Class II molecules have many alleles (point mutations, gene conversion, gene recombination) Diversity ensures that a wide range of peptides can be presented within the population (even if a much more limited set is presented by any individual) For Class II, both a and chains are polymorphic (except DRa in humans and Ea in mice), adding more diversity Collection of Class I + Class II alleles on one chromosome is called the haplotype MHC molecules are expressed co-dominantly: alleles from both chromosomes are expressed in each cell
MHC-Linked Diseases • Defects in MHC gene expression lead to immunodeficiencies (MHC molecules are required for both T cell development and activation) • Some MHC alleles are associated with susceptibility or resistance to autoimmune diseases
MHC-Linked Immunodeficiencies Bare Lymphocyte Syndromes lead to loss of MHC molecule expression: • Defects in TAP genes prevent MHC Class I protein surface expression (even though MHC proteins are normal), so no CD8+ T cells - surprisingly mild immunodeficiency (respiratory and skin infections) • Defects in TF’s controlling Class II gene expression (CIITA, RFXANK, RFX5, RFXAP) block CD4+ T cell development - result in SCID (severe combined immunodeficiency)
Fig. 13.20 Associations of HLA serotype and sex with susceptibility to autoimmune disease.