Download

1 / 40
• 400 likes • 511 Views
Spikes - Exploring the neural code. Kapitel 2. Fred Rieke David Warland Rob de Ryter van Steveninck William Bialek. KAPITEL 1. Spike Spiketrain firing rate wir haben aus Kapitel 1 noch in Erinnerung, dass wir die firing rate nicht allein nutzen

E N D
Spikes - Exploring the neural code Kapitel 2 Fred Rieke David Warland Rob de Ryter van Steveninck William Bialek
KAPITEL 1 • Spike • Spiketrain • firing rate • wir haben aus Kapitel 1 noch in Erinnerung, • dass wir die firing rate nicht allein nutzen • können, um die Signale, welche zu einem • Spiketrain geführt haben, wieder zu • rekonstruieren
Wichtigkeit geringer Spikeanzahl - Das Feldermausgehör : - aus einer Studie von Simmons und Fritz (1993) viele Neuronen zuständig für die Echoverzögerung - auf einen einzelnen Impuls oder einfache Signale, wie normale Töne ist die Reaktion schwach - bei Verzögerung die der ZellenFrequenz entspricht ist die Reaktion genau einem Spike - Visuelles System der Fliege : - eine Fliege ändert ihre Flugbahn innerhalb 30ms, falls eine typische Bewegung ihr visuelles Feld streift - In dieser Zeit können aber nicht viele Spitzen generiert werden
Wichtigkeit geringer Spikeanzahl - Visueller Cortex bei Affen : - Knierem, van Essen (1992) - unterscheidbare Texturen produzieren im Durchschnitt 1-3 Spikes/Zelle innerhalb 50-100ms - Somartosensorischer Cortex bei Ratten : - Fee, Kleinfeld (1994) - Zellen reagieren auf Verschiebung der Schnurr- haare => Entdeckung der Umwelt - es ist sogar wahrscheinlich, dass die Wichtigkeit der geringen Anzahl von Spikes nicht nur für die frühen Phasen der Signalverarbeitung limitiert ist sondern auch darüber hinaus
Wichtigkeit geringer Spikeanzahl - Beispiel : - im visuellen Cortex von Primaten (sehr entfernter Bereich in der Signalverarbeitung) reagieren Zellen sehr wählerisch auf bestimmte Gesichter - Gross, Sergent (1992) - eine 20ms Präsentation ein ~400ms andauerndes Feuern hervorgeruft - wenn aber eine Maske dem Gesicht folgt, so ist der maximale Output in der Regel 5 Spitzen - reicht aber, um Gesicht nachhaltig zu erkennen - überraschender Weise werden sehr viel anspruchsvollere visuelle Aufgaben so schnell ausgeführt, dass jede Ver- arbeitungsschicht genau einen Spike feuern kann, ...
Wichtigkeit geringer Spikeanzahl - ... bevor das „Resultats“-Signal zur nächsten Schicht des Prozesses gelangt - Thorpe, Fize, Marlot (1996) brain storming-Abschnitt : - obwohl geringe Anzahl von Spikes pro Zelle, gibt es nun sehr viele davon - eine Einschätzung über die firing rate, Intervall- verteilung, ... könnte man erhalten durch das Bilden des Durchschnitts über alle Zellen - ganz bestimmt : viele sensorische Signale werden gemeinsam von einer grossen Anzahl von Zellen geteilt (das soll uns nicht entschuldigen über einzelne Spikes nachzudenken)
Wichtigkeit geringer Spikeanzahl - beispielsweise können wir Menschen, wenn wir in einem fast dunklen Raum sitzen, einzelne Photonen sehen Fundamentales Problem : - Annahme das das Nervensystem Zugang zu vielen Spikes einfach deswegen hat, da es soviele Neuronen besitzt - möchte man bspw. die firing rate bestimmen beim Zusammenfassen der Neuronen, die denselben spiketrain besitzen, so muss man die Hypothese machen, dass diese Neuronen dieselben Signale erhalten haben - nur so würde das Zusammenfassen Sinn machen - ...
Wichtigkeit geringer Spikeanzahl Fundamentales Problem(2) : - ... eine weitere Annahme wäre, das diese Reaktionen statistisch unabhängig sind - und die Durchschnittbildung die Zuverlässigkeit des Signals verbessert - wenn diese beiden Hypothesen korrekt sind, so hieße dies, das der Durchschnitt über die Neuronen gleich dem Durchschnitt über die vielen verschiedenen Präsen- tationen derselben Erregung ist - dem entgegen steht ein direkter Beweis (ergibt sich aus den nächsten Abschnitten)
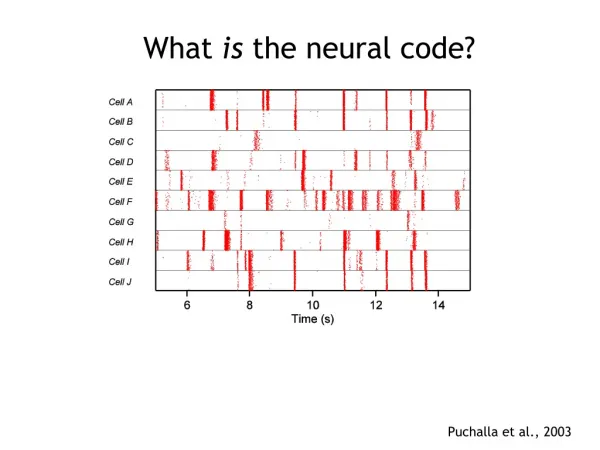
Was kann eine geringe Anzahl von Spikes dem Gehirn mitteilen? - traditionell : durchschnittliches Verhalten der Neuronen als Antwort auf die Veränderung eines Reizes in seinen Parametern wurde katalogisiert - was auch immer die ultimative Charakterisierung des neuralen Codes ist : „es wäre unvernünftig sich vorzustel- len, dass das Auftreten einer einzelnen Spitze zu einer kompletten Gewißheit über die Natur des sensorischen Reizes führt“ - wir brauchen erst einmal eine Sprache, die unser Maß an Gewißheit oder Ungewißheit quantifiziert
Wahrscheinlichkeitstheorie - die konventionelle Annäherung (Kapitel 2.1) an den neuronalen Code kann als Aufteilung der Spikeverteilung in einzelne Scheiben gedacht werden, die die Reaktion zu einem bestimmten Reiz sind : P[{ti} | s()] Wahrscheinlichkeit, dass Spiketrain {ti} generiert wird, wenn Reiz s() stimuliert.
Wahrscheinlichkeitstheorie Sicht des Organismusses : - was weiß man über den Reiz, aufgrund des Spiketrains P[s() | {ti}] Bedingte Verteilung, die die relative Wahrscheinlichkeit von unterschiedlichen Reizwellenformen s() durch den bestimmten (bekannten) spiketrain {ti} mißt.
Wahrscheinlichkeitstheorie - zwei Wahrscheinlichkeitsverteilungen : 1 : die Verteilung der Spiketrains die den Reiz geben 2 : die Verteilung des Reizes, die der Spiketrain liefert - Bayes-Regeln - wir stellen uns vor, alles verstanden zu haben, eingeschlos- sen all der Komplexität des Rauschens, der Adaption und Nichtlinearität - die Verteilung P[{ti} | s()] ist also für alle fixiert - die fundamentale Konsequenz aus den Bayes-Regeln ist nun, dass selbst das komplette Wissen nicht ausreichen würde, um uns zu erzählen was ein gegebener Spiketrain bedeutet und wofür er steht im Bezug auf die äußere Welt!
Wahrscheinlichkeitstheorie - wir beobachten nun eine Sequenz von Spikes zu den Zeitpunkten t1..tn und fragen uns welcher sensorische Reiz diese Spitzen vervorgerufen hat so schauen wir auf folgende Formel : P[s() | {ti}] =P[{ti} | s()]*(1 / P[{ti}]) (2.22) - der erste Term : ist die Verschlüsselungsverteilung (encoding distribution) - der zweite Term : ist die Verteilung der Signale in der Welt - der dritte Term : ist die Wahrscheinlichkeit, genau diese Spikesequenz zu beobachten
Was kann eine geringe Anzahl von Spikes dem Gehirn mitteilen? - auch wenn wir komplett die Verschlüsselung der Signale in Spikes charakterisieren - die Interpretation der Spikes (die diese Signale reprä- sentieren) für die äußere Welt hängt von der Charak- teristik der Welt selbst ab - Kontext ist wichtig - Bayes sagt uns, dass die Vorstellung des Kontextes gleichbedeutend relevant mit der Interpretation der Spiketrains eines einzelnen Neurons ist - wichtiger Aspekt zur Annäherung der Entschlüsselung, wie sie in der Metapher des homoculus eingefangen ist, ist, dass der Reiz unbekannt für das Tier ist
Was kann eine geringe Anzahl von Spikes dem Gehirn mitteilen? - in der natürlichen Umwelt oder im Experiment - Reize werden zufällig ausgewählt P[s(t)] - P[s(t)] bestimmt das Ensemble der Reize - Problem bei Experimenten : - einfaches Ensemble z.B. Sinus-Wellen - nun kann in diesen Fällen s(t) perfekt vorhergesagen - Wissen über das Vergangene z.B. s(t‘<t) - Spiketrain unnötig, um Reiz zu kennen - um anzufangen herauszubekommen, was der Spiketrain bedeutet (Signale der äußeren Umwelt), müssen wir ein Signal aus einem Ensemble auswählen, das groß genug ist - Grund : wir wollen lernen was die äußere Umwelt mit der Beobachtung des Spiketrains zu tun hat
Was kann eine geringe Anzahl von Spikes dem Gehirn mitteilen? - das ist offentsichtlich ein komplett natürlicher Reiz wie z.B. eine Tonbandaufnahme eines Abends am Frosch- teich; dieses ist aber sehr kompliziert => mehr dazu in Kapitel 5.2 - Problem : - bei der Frage, welche Bedeutung hat ein Spiketrain in einem Reizensemble, kann es in jedem Fall zu einer anderen Antwort führen - Spiketrains sind nicht eindeutig!
Antwort-bedingte Ensembles - es ist möglich geworden eine experimentelle Charak- terisierung der bedingten Verteilung P[s(t) | {ti}] (von einer Antwort eines bewegungssensiblen Neurons) im visuellen System einer Fliege zu bestimmen - de Ryter van Steveninck, Bialek (1988) kurzer Überblick über das Sehen einer Fliege und die Rolle der visuellen Bewegungsabschätzung in dem Fliegenverhalten : - Flugbahn einer Fliege im Raum mit geraden Segmenten und scharfen Richtungswechseln - wenn Licht aus, dann landet die Fliege - Analyse ergab : Richtungswechsel mit 30ms Verzögerung
Antwort-bedingte Ensembles - man kann den Input der Flugkontrolle demonstrieren : - Fliege wird angebunden (hängt mit schlagenden Flügeln) - das visuelle Umfeld der Fliege wird gedreht, nun fängt die Fliege an sich ebenfalls zu drehen - versucht die Rotationsbewegung auszugleichen - man darf aber nicht vergessen, dass ein Luftzug, die Flügelhärte, sowie andere Faktoren ebenfalls Einfluß auf die Flugbahn nehmen - nicht allein eine einzige Reaktion (kein Autopilot) - Fliege lernt sich immer wieder anzupassen - die Verteilung P[s(t) | {ti}] liefert als Effekt, ein Wörterbuch für den neuronalen Code, in dem wir nach den Reizen suchen die am wahrscheinlichsten sind
Antwort-bedingte Ensembles - wir bestimmen also manche Klassen von Reizen als mehr wahrscheinlich als andere, aufgrund der Beobachtung eines bestimmten Spiketrains {ti} - hier tut Erklärung Not...
Lesen des Codes - Grundfrage : Was können wir bei gegebenem Spiketrain über die unbekannte Reizwellenform s(t) sagen? - eine mögliche Antwort : - wir können den Spiketrain „lesen“ und ständig zum Reiz zurückübersetzen - sehr einfache Antwort (wenn richtig) - eignet sich für quantitative Analyse : - Wie genau sind die konstruierten Wellenformen? - Wie komplex ist die Rekonstruktion der Algorithmen? - Wie beeinflussen Fehler der Messung der Spikean- kunftszeit die Rekonstruktion? - Reizrekonstruktion wird nicht notwendigerweise vom Tier gelöst
Lesen des Codes - Charakter aber der gleiche - Beispiel : - Fliege kann Wende einleiten, die nur auf visuelle Bewe- gungssignale basiert - bedeutet : Spikeoutput des Visualneurons wird in Dreh- moment übersetzt - Signal des Drehmoments ist kontinuierliche Wellenform - das Problem der Wiedergewinnung analoger Signale aus dem Spiketrain ist dann eine fundamentale Stufe in der neuronalen Verarbeitung der Sensor-Daten
Warum kann es funktionieren - Hauptschwierigkeiten : - Spärlichkeit, Zufälligkeit - wir benötigen Lernen zur Decodierung um Reiz-Wellen- form zu schätzen - Viele verschiedene Reize können denselben Spiketrain produzieren und wiederholte Reizpräsentationen produ- zieren nicht identische Spiketrains - Frage : Ist Decodierung dann überhaupt möglich? - wenn P[s(t) | {ti}] einen spitzen Peak an einer bestimmten Reiz-Wellenform s°(t; {ti}) darstellt, ergibt es einen Sinn zu sagen, dass der Spiketrain {ti} „für diesen Reiz steht“ - Breite des Peaks misst Genauigkeit mit der sich s°(t; {ti}) dem wahren Reiz nähert
Lesen des Codes - wenn P[s(t) | {ti}] ohne deutlichen Peak breit verschmiert ist; keinen Sinn für Wiedergewinnung - dazwischen P[s(t) | {ti}] hat mehrere Peaks oder Maxima- Reihe; mehrdeutig Abb.2.17 a) gut aufgelöster Peak gute Decodierung b) & c) keinen klar erkenn- baren Peak
Lesen des Codes - Bialek, Zee (1990) - motiviert durch experimentelle Ergebnisse bei Fliegen - formulierten das Decodierungsproblem im Kontext einfacher Modelle für die Statistik der Spikecodierung (insbesondere Poissonmodell aus 2.1.4) - es stellt sich heraus, dass das Problem der Reizschätzung s(t) bei gegebenem Spiketrain {ti}, dem Problem der Vor- hersage der Trajektorie eines Teilchens zu einem Impuls, immer wenn ein Spike erscheint, äquivalent ist - wenn es wirklich möglich ist den Reiz aus dem Spiketrain zu rekonstruieren, dann diese Kombination aus gleichför- migen und impulsartigen Kräften verursachen, dass sich die Trajektorien um die wahre Reiz-Wellenform zusammen- ballen
Lesen des Codes - da Verbindung zwischen Spikes und Signalen wahrscheinlich ist, müssen wir Rekonstruktionsproblem präziser definieren - natürliche Wahl : - Frage nach dem wahrscheinlichsten Reiz oder äquivalent, die wahrscheinlichste Trajektorie bei gegebener Kraft - maximum Likelihood-Schätzung - andere natürliche Wahl : - mittlere Reizwellenform berechnen - optimal, da der mittlere quadratische Fehler zwischen geschätztem und wahrem Reiz minimiert wird - beide ähnliche Ergebnisse - nun legen wir die Konzentration auf bedingtem Mittelwert
Lesen des Codes - aus Diskussion 2.1.3 wissen wir, dass in vielen Systemen die mittlere Trajektorie eines Teilchens in linearer Be- ziehung zu den angewandten Kräften steht : <x(t)> = K1() F(t - ) d (2.23) - K() die lineare Reaktionsfunktion (Fouriertransformierte?) - Annahme : mittlere Position in Abwesenheit einer Kraft ist Null - in Analogie zur Spike-Codierung besteht Kraft aus einer Reihe von Impulsen zur Spikezeit ti F(t) = i=1;n (t - ti) (2.24)
Lesen des Codes - und die Trajektorie x(t) stimmt mit der Signal-Wellenform s(t) überein - somit stimmt die lineare Reaktion des mechanischen Sys- tems mit der linearen Rekonstruktion des Signals aus dem Spiketrain überein : sest(t) = K1() i=1;n (t - - ti) d = i=1;n K1(t - ti) (2.25) - Bedeutung (2.25) : - anstatt zu charakterisieren, wie Nervensystem Signale in Spikes wandelt, char. (2.25) den Vorgang, durch den ein Beobachter des Spiketrains den Reiz schätzen oder re- konstruieren könnte - Black-Box aus Abbildung 2.18 annähernd eine lineare Einrichtung
Lesen des Codes Abb.2.18 Schematik der Reiz estimation - allgemeiner : wir erlauben der „Black-Box“ Nichtlinearitäten zu haben, das stimmt mit der Tatsache überein, dass das Teilchen beim äquivalenten Problem der statistischen Mechanik, eine nichtlineare Reaktion auf die angewandten Kräfte zeigt : sest(t) = <s(t)> = i K1(t - ti) + 1/2 * i K2(t - ti, t - tj) + ... (2.26)
Lesen des Codes - bei der Analyse eines Modell-Neurons ist es möglich alle Kerne K1, K2, ... zu berechnen - tatsächlich hängt die Struktur der Kerne von der Statistik der Input-Signale ab, so dass die optimalen Strategien zur Lesung des neuronalen Codes, von der Natur des Signals in der Umwelt abhängt - Kn(t) keine Eigenschaft des Neurons allein, sondern viel- mehr Eigenschaften des Neurons und seiner sensorischen Umwelt vereint - Bialek, Zee (1990); Gabbiani, Koch (1996)
Lesen des Codes - jetzt kamen ca. 6 Seiten, um folgende Aussage zu veri- fizieren : - Kolmogorov-Wiener-Filter ist ein linearer Filter, der jedes lineare Schätzungsproblem optimal löst 2.3.2 Eine experimentelle Strategie - Problem des Lesens des neuronalen Codes als die Konstruktion eines (allgemein linearen) Filterns formuliert, der die Spikes als Input behandelt und eine Schätzung des sensorischen Reizes wie in Gl. (2.26) produziert - Ziel nun : empirische Näherung finden, um bei geg. s(t) und dem gem. {ti}, die Kerne K1(), K2(), ... auszuwählen
Lesen des Codes - wir brauchen noch eine Fehlerfunktion, die unsere Schätzung beschreibt E[s(t) , sest(t)] Quadratische Fehlerfunktion - Kausalität - zeigt uns, dass ein Spike unsere Reizschätzung nicht be- einflussen kann, bevor der Spike erschienen ist - Kerne für negative Zeiten muss NULL sein - bedeutet nicht, dass ein Spike aus der Vergangenheit unsere Schätzung nicht beeinflussen kann - strenge kausale Beziehung zwischen Reiz und SpikeZeiten
Lesen des Codes - Problem : ein Spike beeinflusst unsere Reizschätzung nur, nachdem der Spike aufgetreten ist, aber der Reiz beeinflusst die Bildung des Spikes nur, bevor der Spike auftritt - Lösung : wir akzeptieren eine Verzögerung bei der Schät- zung; Grösse hängt von Code-Struktur ab - nun haben wir ein gut mathematisch formuliertes Problem : - finden die Kerne Kn, die die Fehlermessung minimieren und dabei dem Zwang der Kausalität gehorchen (Probleme können noch bei lokalen Minimas auftreten) - Bei unseren Experimenten tritt dieser Fall jedoch nicht auf 1. Fehlermessung ist quadratisch (keine lokalen Minima) 2. Datenmenge ziemlich gross (10 „hoch“ 5 Spikes)
Lesen des Codes 2.3.3 Qualitative Merkmale eines ersten Tests Abb.2.20 a) Schätzungsfilter (aus Exp.) Spikereaktionen auf kurzen Teil des Reizes (punkt. Kurve in (c) sind in (b) gezeigt). Aufgerichte- te Spikes sind Reaktionen auf Reiz. Abwärts auf denselben nur andere Richtung (2 Zellen). Die Schätzung ist durchgezogene Kurve in (c) mit Gaußfilter geglät- tet (5ms)
Lesen des Codes - Abbildung zeigt, dass wir, auch wenn H1 nichtlinear reagiert, die Spikes mit einem linearen Filter decodieren können : Abb.2.22 hier wurde mittlerer Reiz aus einem gegebenen bestimmten Rekonstruk- tions-Wert gegen den Rekonstruk- tions-Wert selbst aufgetragen Systematische Fehler in der Schät- zung sollten sich als Abweichungen darstellen. Das Fehlen weist darauf hin, dass nichtlineare Terme nichts signifikant zum Schätzungsprozess beitragen.
Lesen des Codes - eine andere Art, den Erfolg einer linearen Rekonstruktion zu prüfen, wäre in die Gl. (2.26) weitere Terme einzufügen, und zu schauen, ob die Qualität verbessert werden kann - kurze Antwort : es gibt keine statistisch, signifikanten Veränderungen - vielleicht war die quadratische Fehlermessung zu grob für die Qualitätsprüfung (wird diskutiert in 3.2.3) - Fliege ist mit einem interessanten Dilemma konfrontiert, das vielleicht für die Sensor-Signal-Verarbeitung typisch ist. - Verhaltensbetrachtungen drängen auf kurze Entscheidungs- zeiten; doch kurze Ent.Zeiten bedeuten, dass das System in jeder Verarbeitungsstufe für das Rauschen anfällig ist
Lesen des Codes - um die Relation zwischen Zuverlässigkeit und Entscheidungs- zeiten zu untersuchen, kann die kausale Rekonstruktion bei verschiedenen Verzögerungszeiten gefunden werden - um Qualität dieser unterschiedlichen Rekonstruktionen zu testen, berechnen wir die Kreuzkorellation der Rekonstruk- tionen mit dem Reiz : Abb.2.23 normalisierte Kreuzkorellation zwischen Reiz und Rekonstruktion für versch. Verzö- gerungen; ebenfalls gezeigt Autokor. des Reizes (Kurve s), die mit Gauß-Filter mit einer Std-Abweichung von 5ms geschätzt wurde. Die Kreuzrelation steigt zw. 10-40ms an. Weitere Anstiege bei der Verzögerung haben minimalen Effekt.
Lesen des Codes - wir haben die Reaktion der Zellen in einer einfachen Welt beschrieben, inder nur die Winkelgeschwindigkeit in der Zeit variiert - tatsächlich hängt die Feuerrate von der räumlichen Struktur ab - Was geschieht, wenn wir die räumliche Umwelt verändern? - Genauigkeitschwankungen werden in 4.3.3 behandelt
Zusammenfassung Es wurde eine Näherung zur Neural-Codierung vorgeschlagen, die sich auf das Verhältnis konzentriert, das ein Tier aus seinen eigenen neuralen Signalen über die sensorische Umwelt ableiten kann, und darauf, wie diese Information aus den neuralen Reaktionen herausgezogen werden kann. Dies ist ein Problem, das vom Homunculus gelöst werden muss. Das Problem Spiketrains zu interpretieren, scheint eine ein- fache Lösung zu haben, mit der es möglich ist, die Wellenform unbekannter Reize aus Beobachtungen eines einzelnen Spike- trains, direkt zu schätzen. Das Schätzungsverfahren ist sehr einfach und besteht im wesentlichen aus einem geeignet ge- wählten linearen Filter. Obwohl die Details dieses Filters wahrscheinlich nicht wichtig sind, erlaubt uns seine Struktur, zu sehen, wie der Code passend zum Verhalten ist.
Fazit zum Vortrag Übersetzungsprobleme wurden gemeistert von : - Michael Schreiber - Günter Pehl Text ist für einen Laien sehr schwer zu lesen und kaum zu verstehen. Für eine Seminararbeit war das zu viel! Zu anspruchsvolles Thema, verpackt in einer Fremdsprache. Vielen Dank für‘s zuhören...