Download

1 / 57
580 likes | 787 Views
インタラクトームとしての Dynamical Networks 我らオーム主義者なり (K.Nakai). 鈴木泰博、荻島創一、長谷武、中川草、田中博 東京医科歯科大学 大学院疾患生命科学研究部 生命情報学 ( ATR は今年度(平成 15 年)末まで). 注意! 私は 医者ではありません。 ましてや 歯医者でもありません。 講演後に医療相談にはのれません。. 私は 生物学者でもありません 。 小難しい横文字の言葉をちりばめて 質問して苛めるのはやめてください。 わかっている“フリ” … してるだけです。 不勉強です。浅学です。 温かい目で見てあげてください。

E N D
インタラクトームとしてのDynamical Networks我らオーム主義者なり (K.Nakai) 鈴木泰博、荻島創一、長谷武、中川草、田中博 東京医科歯科大学 大学院疾患生命科学研究部 生命情報学 (ATRは今年度(平成15年)末まで)
注意! 私は医者ではありません。 ましてや歯医者でもありません。 講演後に医療相談にはのれません。 私は生物学者でもありません。 小難しい横文字の言葉をちりばめて 質問して苛めるのはやめてください。 わかっている“フリ”…してるだけです。 不勉強です。浅学です。 温かい目で見てあげてください。 ごめんなさい。 鈴木泰博(るろうの たみ) 初手からして理論計算機科学でドロップアウト その後、常にアウトからアウトへ、左から左へと放浪 経営学という遠い島から流れ来る“椰子の実研究者”
Back ground 1 What’s DNA and protein? Back ground 2 我らオーム主義者なりby Nakei.Kbio-networks as interactome Quick View Dynamical networks と分子生物学 出芽酵母のタンパク相互作用ネットワークについて 最後に(創薬・医療とinteractome) Agenda
はじまりのはじまり 地球上の生命体はすべて同じ種類のタンパク質でできています。 これは、当たり前に思えるかもしれませんが、ものすごい特徴です。 なぜなら、 これを逆手にとれば、タンパク質をうまくデザインすれば生命体ができるからです ゲノムにはタンパク質をどのようにデザインすれば良いか が書いてあるのです
DNAそれはバイブルのバイブル DNA 遺伝子 エクソン イントロン イントロン sidafdmorearegwatashinonamaehasuzukidesuremoreaofgamfeoafmeoafea 翻 訳 タンパク質 アルファベット 24文字 わたしのなまえはすずきです すべての生物は同じ“言語”で 書かれたDNAを持ち、 同じ種類のタンパク質の組み合わせ でできています。 ひらがな50文字 A,U,C,T 4文字 タンパク質 20種類
複数の遺伝子間ではネットワークを形成する 反比例 negative feed back wacmeogashitasikencacdacashitadetocdacamoashitahahayaokicacdamoeacamdoa 正比例 positive feed back スイッチON! wacmeogashitasikencacdacashitadetocdacamoashitahahayaokicacdamoeacadoa 明日試験 明日デート 明日早起き スイッチON! wacmeogashitasikencacdacashitadetocdacamoashitahahayaokicacdamoeacamdoa 明日デート 明日早起き 明日試験
遺伝子からタンパク質へそして生命体としての機能へ遺伝子からタンパク質へそして生命体としての機能へ タンパク質が機能するためには、それなりの条件が必要です。 それを人工的につくるのは容易ではありませんが、生体ではそれをきちんと機能させているのです。
“やんごとなき”たんぱく質の生涯揺り篭から墓場まで“やんごとなき”たんぱく質の生涯揺り篭から墓場まで Anfinsenのドグマ
社会システムとしての細胞 枠組みを与えるDNA 実質的に社会を構成するプロテイン群 バイブルとしてのDNA 実態を支えているのはタンパク質
Back ground 1 What’s DNA and protein? Back ground 2 我らオーム主義者なりby Nakei.Kbio-networks as interactome Quick View Dynamical networks と分子生物学 出芽酵母のタンパク相互作用ネットワークについて 最後に(創薬・医療とinteractome) Agenda
Interactomeオーム主義の時代 オーム主義万歳 ゲノ(遺伝子)+オーム プロテ(タンパク)+オーム トランスクリプト(転写)+オーム シグナロソ(シグナル伝達)+オーム インタラクト(相互作用ネットワーク)+オーム(集団・全体) 最近では“ネットワーク生物学”なぞと云うコトバまで。。 厳密な定義?。。。知りません
OVER VIEWinteract-ome (biology] プロテオーム MASベース 比較的小規模 ネットワーク モチーフの提唱 • 生物ネットワークのスクリーニング • タンパク相互作用大規模スクリーニング • (Y2H, TAP, HMS-PCI ) • 出芽酵母(2000-), • pyroly(2001), • C.elegence (2003), • ショウジョウバエ(2003), • 大腸菌(2004)予定 • 遺伝子ネットワーク • 出芽酵母(2004) 臨床・創薬への応用 我々の目的 機能予測 出芽酵母(2000-) Comparative netwroks 出芽酵母タンパク相互作用(2000) 出芽酵母遺伝子ネットワーク(2004) 応用 基礎
PNAS2001 他に、Utez, Tongらのフルスクリーニングがあるデータベースとしては YPD, MIPS, SGDなどがある。
H.Pyloriのタンパク相互作用ネットワーク 2001 Nature
flyDrosophia melanogaster 2003 Science C.elegence Multi cellular function related protein のPPI はスクリーニング完了(2004 Science)
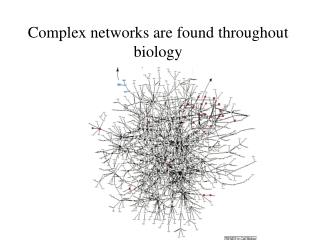
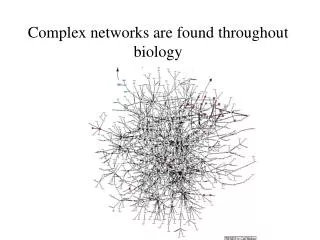
classic①A network of protein-protein interactions in yeast (Schwikowski et al. 2000) MIPS 1183 YPD 471 (2,358 interactions) Screening DIP 76 1,548 proteins/2,358 links Ito 134 Utez 808 each number indicates the number of interactions not the total number of stored proteins 19 proteins 5-11 proteins (9 networks) less than 4 (193 networks)
A network of protein-protein interactions in yeast (Schwikowski et al. 2000) MIPS 1183 each node and color stands for a protein a function. YPD 471 (2,358 interactions) Screening DIP 76 Ito 134 Protein A—Protein B Rank Function total 1. Cromatin struc. 5 2. memb. Fusion 3 3. cell struc. 2 4. lipid memb. 1 4. endcyt. 1 Utez 808 each number indicates the number of interactions not the total number of stored proteins This protein has and functions. 72% can cover Based on YPD 42 cellular role; a protein engaged in and not its precise biochemical activity (Utez, Tong)
A network of protein-protein interactions in yeast (Schwikowski et al. 2000) YPD 2,039 proteins 554 have no annotation prediction prediction 364/554 have at least one partner of known func. 69/364 have two or more partner of known func. 29/69 have two or more partner of known func. in common Marcotte et al. annotate proteins in yeast by using as much as possible kind of method Y2H, phase display, (science 285 1999)
最近の流れ 複数のタンパク間で“共有”するタンパク質の割合で機能予測を行う (Samanta, Liang, Vazquez) 特性が似ているならば同様のタンパクと相互作用するであろう さらに関係に機械学習を導入したものもある(阿久津ら 2003)
2002 Nature 新たなスクリーニングの方法 (HMS-PCI法の応用) High-throghput mass spectrometric protein complex identification他にTAP法などが応用されるようになってきた。 R;unknown B:known
インタラクトームclassic② 2002 Science 配列情報とY2Hの実験結果を組み合わせ、結合する塩基配列を 予測する方法を確立し、その方法を用いて出芽酵母のSH3ドメイン 結合性タンパクの相互作用解析に応用。 グラフ理論の応用により、SH3ドメイン関連のタンパク質内に疾患 関連タンパクネットワーク(WASP症候群)が存在することを確認。 この結果はin vivoの結果とも一致をみた。 Wiskott-Aldrich症候群(WASP)病因:X連鎖遺伝形式をとり、血小板減少と湿疹を合併する免疫不全症である。細胞内骨格の機能やシグナル伝達に関わるとされるWASP遺伝子の異常による。X連鎖血小板減少症は同じWASP遺伝子異常による。臨床症状:血小板減少による出血傾向とアトピー性皮膚炎要の湿疹があり、細菌、真菌、ウイルスに対する易感染性を認める。
相互作用の最適化制御 2003 Nature タンパクはなにかと絡みつきやすいもので。。。 出芽酵母のSH3ドメインにおいて Pbs2キナーゼ モチーフのあるモチーフが相互作用を最適化して いる。ここで、最適化とはシグナル応答の際に競合 するプロテインの相互作用をブロックして、円滑に必 要な相互作用が行われるようにすることを意味する。 Pbs2のこの活性はSH3ドメイン以外ではみられない。
システム生物学的な潮流 Nature 2002
Back ground 1 What’s DNA and protein? Back ground 2 我らオーム主義者なりby Nakei.Kbio-networks as interactome Quick View Dynamical networks と分子生物学 出芽酵母のタンパク相互作用ネットワークについて 最後に(創薬・医療とinteractome) Agenda
OVER VIEWineract-ome [数理&biol.] 臨床・創薬への応用 我々の目的 シグナル伝達系 の特徴 classic Jeong, Hub proteins と致死性 Y2Hデータの純化 (斉藤、中谷) 統計的性質 ネットワークの進化 G.Wagner などなど 数理 生物
classic致死性タンパクと相互作用との関係Jeong 2001, Nature • 出芽酵母のタンパク相互作用ネットワークにScale Free性を確認 (pyrolyについても…他論文) • 相互作用の多い(hub)タンパクが致死性と高い相関があることを確認
遺伝子ネットワークのスクリーニング Science 2004 SF, small world 性を有する
グラフ理論の応用 中谷:ネットワークの比較によるタンパクー遺伝子間の関係の推定 他に、斉藤のY2Hデータの純化の研究などが知られている
プロテインネットワークの進化 しかし、大腸菌、白イナズナ、分裂酵母、出芽酵母 の比較を行うと、hubとなっているタンパクが進化速 度が遅いとはいえない。 進化的に古いタンパクがhubプロテインではないこと が示されている (koonin) タンパクネットワークの進化はPreferential Attachment ではないことが示されている。 G.Wagnerらは精力的に遺伝子重複の 数理モデル的解析を行っている。
2000Nature ネットワークのアキレス腱としてのP53
DNA damage p53 p53 degradation Signaling Network Preliminary model input Mdm2 tetrameri -zation p53 p53 tetramer p53 p53 activation p53 translocation p53 activated tetramer Mdm2 induction input p53 nucleus G1 arrest DNA repair cytoplasm cytoplasm p53, mdm2 -> empty p53 -> p53[n] nucleus P53,p53,p53,p53 -> p53_tetramer p53_tetramer -> p53_tetramer, Mdm2 p53, Mdm2 -> empty Mdm2 -> Mdm2[c] p53_tetramer, Mdm2 -> p53,p53,p53,p53[c] p53_tetramer, DNA damage -> p53_tetramer+, DNA damage p53_tetramer+, DNA damage -> p53_tetramer WHERER, they assign translocation rule [n] nucleus import [c] nucleus export
無理やりまとめてみると。。 • 背景 • ① 生物ネットワークの大規模スクリーニングデータが近年になって増加してきた。 • ② Dynamical Networksの研究の進展 • 現在の方向性としては、 • Bioinformatics; タンパクの機能の推定アルゴリズム、Y2Hデータの純化 • Proteomics;比較ネットワーク • 分子進化
だいぶ、講演時間を 長くするために 水増ししてましたけど やっと次から 研究の話です。 生物学者 たちとの 語らい
Agenda • Back ground 1 What’s DNA and protein? • Back ground 2 我らオーム主義者なりby Nakei.Kbio-networks as interactome • Quick View Dynamical networks と分子生物学 • 出芽酵母のタンパク相互作用ネットワークについて • 最後に(創薬・医療とinteractome)
レポート課題ネットワーク生物学 レポート問題:タンパク相互作用ネットワークのデータを用いてその特徴について調べなさい。ただし、タンパク相互作用のデータはMIPS, YPD, ITO, Utez, Tongのどれを用いてもよい。 提出期限:特に定めませんが、誰かが先にやっていた場合はそのレポートは零点となります。 提出先:いずれかの国際誌(ただし、査読をパスする必要があります)。
従来の関連研究Specificity in topology in the yeast protein network (Maslov, Science, 2002) The higher degree nodes are liable to be connected to the lower degree nodes Results from Y2H keep degree the same Random shuffling compare the change of connectivity H->L Null model との比較
Yeast 2 Hybrid system (Y2H) prey z bait 方向に意味なし x A→Bならば、B→Aとなるべき であるが、Y2Hではそうならない 場合が多い。 1.両方向を確認した場合。(Tong) 2.1.でない場合は、ベクタ内の prey proteinsがbaitに結合するか は、確率的なので双方向にならない とは断定できない。 Y2Hでは対称性が崩れる場合が多いがそれは意味のある事なのか、単なる実験の特性なの かよくわからない。
Bait-prey の非対称性①(出芽酵母) outgoing bait incoming prey
WHYOUT-IN = 1? (やったこと①) Out degree , In degree Out-Indegree の頻度分布 1 frequency 出芽酵母でもピロリ菌でも OUT-IN=1 となるプロテインが多い。 Out-Indegree
シミュレーション(初期分布の設定) Out-In=1…で結合のノードの分布を生成 分布 P(k)=k-2 で結合の分布を生成 Out-In degree の頻度分布 Out+In degree の頻度分布 frequency frequency Out+In degree Out-In degree OUT-IN の頻度分布を計測
結合後のOut-In degreeの分布 中心=5 中心=1 1 1 frequency frequency Out-Indegree OutーIndegree 中心=10 中心=30 1 1 frequency frequency Out-In degree Out-In degree
機能と非対象性との関係(やったことその②)機能と非対象性との関係(やったことその②) 機能数 機能数 OUT-IN OUT+IN
Correlation between the number of proteins and functions 多機能のタンパクは高い相互作用を持つ many functions produced byfewproteins 40functions/2proteins 40/2=20.0 functions/protein 40functions/200proteins 40/200=0.5 functions/protein
非対称性と機能数の関係 Out+In 10 8 平均機能数 Out-In 6 Ave. functions 4 2 outgoing+incoming 0 outgoing-incoming 0 10 20 30 Outgoing+/-incoming
(11 1) ENERGY_GENERATION SMALL_MOLECULE_TRANSPORT CELL_CYCLE_CONTROL PROTEIN_COMPLEX_ASSEMBLY CYTOKINESIS MITOSIS DNA_SYNTHESIS CHROMATIN_OR_CHROMOSOME_STRUCTURE RNA_SPLICING RNA_PROCESSING_OR_MODIFICATION POL_II_TRANSCRIPTION OTHER_METABOLISM MEIOSIS PROTEIN_DEGRADATION VESICULAR_TRANSPORT DNA_REPAIR (8 2) SMALL_MOLECULE_TRANSPORT MATING_RESPONSE PROTEIN_SYNTHESIS CELL_STRESS CELL_POLARITY SIGNAL_TRANSDUCTION RNA_SPLICING RNA_PROCESSING_OR_MODIFICATION (6 2) OTHER_METABOLISM (10 2) MEMBRANE_FUSION VESICULAR_TRANSPORT RECOMBINATION MEIOSIS DIFFERENTIATION CELL_STRESS CELL_POLARITY (8 3) SIGNAL_TRANSDUCTION CELL_STRESS (4 1) PROTEIN_DEGRADATION CHROMATIN_OR_CHROMOSOME_STRUCTURE PROTEIN_MODIFICATION SMALL_MOLECULE_TRANSPORT AMINO_ACID_METABOLISM POL_II_TRANSCRIPTION CELL_CYCLE_CONTROL (15 3) MITOSIS PROTEIN_DEGRADATION MATING_RESPONSE CELL_CYCLE_CONTROL PROTEIN_SYNTHESIS AMINO_ACID_METABOLISM CELL_POLARITY (12 1) PROTEIN_COMPLEX_ASSEMBLY PROTEIN_TRANSLOCATION SMALL_MOLECULE_TRANSPORT LIPID_FATTY_ACID_AND_STEROL_METABOLISM VESICULAR_TRANSPORT (1 0) PROTEIN_MODIFICATION OTHER_METABOLISM CHROMATIN_OR_CHROMOSOME_STRUCTURE RNA_PROCESSING_OR_MODIFICATION SMALL_MOLECULE_TRANSPORT DNA_SYNTHESIS POL_II_TRANSCRIPTION CARBOHYDRATE_METABOLISM (4 0) PROTEIN_DEGRADATION CHROMATIN_OR_CHROMOSOME_STRUCTURE MITOSIS AMINO_ACID_METABOLISM CELL_CYCLE_CONTROL MITOSIS CELL_STRESS CARBOHYDRATE_METABOLISM MEIOSIS PROTEIN_SYNTHESIS POL_III_TRANSCRIPTION POL_I_TRANSCRIPTION POL_II_TRANSCRIPTION MEIOSIS CARBOHYDRATE_METABOLISM RNA_PROCESSING_OR_MODIFICATION RNA_SPLICING DNA_REPAIR NUCLEOTIDE_METABOLISM CELL_STRUCTURE VESICULAR_TRANSPORT POL_II_TRANSCRIPTION PROTEIN_SYNTHESIS PROTEIN_SYNTHESIS
これらのうち Complex形成 に関与するもの 細胞内局在で分けたインタラクションとコンプレックス形成 Y2H(Network) 局在データなし この テーブル そのうち、WONら に局在が確かめら れた。ネットワーク WON-KI, Nature 局在のデータはWON-KI HUH, et al ”Global analysis of protein localization in budding yeast” Nature 16 Oct 2003 686-691 Complex dataはMIPSのcomplex.171103 より
クリーク構造は生物学的機能と関係がある? タンパク質のPPIネットワーク&アノテーション ミトコンドリア リボゾームタンパク質 タンパク質分解関連 スプライシングに関わる snRNP関連 セリン/スレオニンキナーゼ Translation initiation factor Involved in endoplasmic-to-Golgi protein trafficking 核膜孔を介した 核-細胞質間輸送 アクチン細胞骨格系 DNA修復 密に関連するタンパク質は複合体を形成している傾向がある RNA recognition motif-containing protein Telomere elongation protein (ever shorter telomeres) Glycogen accumulation. Gip1p-Glc7p phosphatase complex is required for proper septin organization and initiation of spore wall formation during sporulation. 寄せ集めの アノテーション で得た結果です May participate in nucleolar function during the transition from stationary phase to rapid growth 機能未知 Probable translation initiation factor 6 (EIF6) similar to mammalian elongin A, interacts with elongin C