Download
1 / 1
10 likes | 131 Views
Effect in CBA Mice of varying Signal Intensity and Age on the Inhibition of the Acoustic Startle Reflex produced by a Change in Sound Location along the Azimuth James R. Ison & Paul D. Allen

E N D
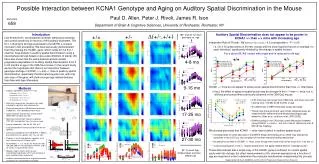
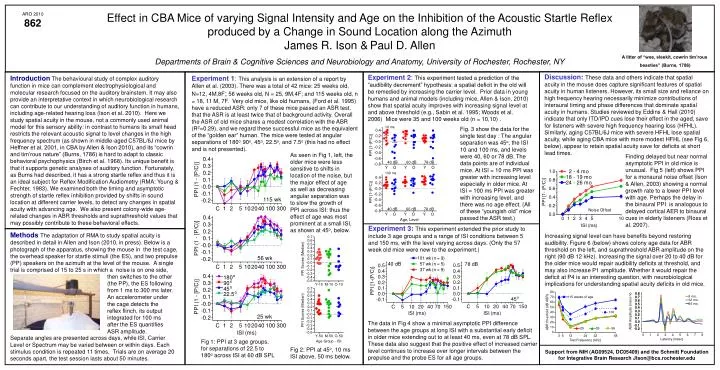
Effect in CBA Mice of varying Signal Intensity and Age on the Inhibition of the Acoustic Startle Reflex produced by a Change in Sound Location along the Azimuth James R. Ison &Paul D. Allen Departments of Brain & Cognitive Sciences and Neurobiology and Anatomy, University of Rochester, Rochester, NY ARO 2010 862 A litter of “wee, sleekit, cowrin tim’rous beasties” (Burns, 1786) Discussion: These data and othersindicate that spatial acuity in the mouse does capture significant features of spatial acuity in human listeners. However, its small size and reliance on high frequency hearing necessarily minimize contributions of interaural timing and phase differences that dominate spatial acuity in humans. Studies reviewed by Eddins & Hall (2010) indicate that only ITD/IPD cues lose their effect in the aged, save for listeners with severe high frequency hearing loss (HFHL). Similarly, aging C57BL/6J mice with severe HFHL lose spatial acuity, while aging CBA mice with more modest HFHL (see Fig 6, below),appear to retain spatial acuity save for deficits at short lead times. Experiment 2: This experiment tested a prediction of the “audibility decrement” hypothesis: a spatial deficit in the old will be remedied by increasing the carrier level. Prior data in young humans and animal models (including mice, Allen & Ison, 2010) show that spatial acuity improves with increasing signal level at and above threshold (e.g., Sabin et al. 1995; Woods et al. 2006) Mice were 35 and 100 weeks old (n = 10,10) . IntroductionThe behavioural study of complex auditory function in mice can complement electrophysiological and molecular research focused on the auditory brainstem. It may also provide an interpretative context in which neurobiological research can contribute to our understanding of auditory function in humans, including age-related hearing loss (Ison et al. 2010). Here we study spatial acuity in the mouse, not a commonly used animal model for this sensory ability: in contrast to humans its small head restricts the relevant acoustic signal to level changes in the high frequency spectrum (as shown in middle-aged C57BL/6J mice by Heffner et al. 2001, in CBA by Allen & Ison 2010), and its “cowrin and tim’rous nature” (Burns, 1786) is hard to adapt to classic behavioral psychophysics (Birch et al. 1968). Its unique benefit is that it supports genetic analyses of auditory function. Fortunately, as Burns had described, it has a strong startle reflex and thus it is an ideal subject for Reflex Modification Audiometry (RMA: Young & Fechter, 1983). We examined both the timing and asymptotic strength of startle reflex inhibition provided by shifts in sound location at different carrier levels, to detect any changes in spatial acuity with advancing age. We also present colony-wide age-related changes in ABR thresholds and suprathreshold values that may possibly contribute to these behavioral effects. Experiment 1:This analysis is an extension of a report by Allen et al. (2003). There was a total of 42 mice: 25 weeks old, N=12, 4M,8F; 56 weeks old, N = 25, 9M,4F; and 115 weeks old, n = 18, 11 M, 7F. Very old mice, like old humans, (Ford et al. 1995) have a reduced ASR: only 7 of these mice passed an ASR test, that the ASR is at least twice that of background activity. Overall the ASR of old mice shares a modest correlation with the ABR (R2=0.29), and we regard these successful mice as the equivalent of the “golden ear” human. The mice were tested at angular separations of 180o, 90o, 45o, 22.5o, and 7.5o (this had no effect and is not presented). Fig. 3 show the data for the single test day : The angular separation was 45o, the ISI 10 and 100 ms, and levels were 40, 60 or 78 dB. The data points are of individual mice. At ISI = 10 ms PPI was greater with increasing level especially in older mice. At ISI = 100 ms PPI was greater with increasing level, and there was no age effect. (All of these “youngish old” mice passed the ASR test.) As seen in Fig 1, left, the older mice were less sensitive to shifts in location of the noise, but the major effect of age as well as decreasing angular separation was to slow the growth of PPI across ISI: thus the effect of age was most prominent at a small ISI, as shown at 45o, below. Finding delayed but near normal asymptotic PPI in old mice is unusual. Fig 5 (left) shows PPI for a monaural noise offset (Ison & Allen, 2003) showing a normal growth rate to a lower PPI level with age. Perhaps the delay in the binaural PPI is analogous to delayed cortical AER to binaural cues in elderly listeners (Ross et al. 2007). Experiment 3: This experiment extended the prior study to include 3 age groups and a range of ISI conditions between 5 and 150 ms, with the level varying across days. (Only the 57 week old mice were new to the experiment.) Methods The adaptation of RMA to study spatial acuity is described in detail in Allen and Ison (2010, in press). Below is a photograph of the apparatus, showing the mouse in the test cage, the overhead speaker for startle stimuli (the ES), and two prepulse (PP) speakers on the azimuth at the level of the mouse. A single trial is comprised of 15 to 25 s in which a noise is on one side, Increasing signal level can have benefits beyond restoring audibility. Figure 6 (below) shows colony age data for ABR threshold on the left, and suprathreshold ABR amplitude on the right (90 dB 12 kHz). Increasing the signal over 20 to 40 dB for the older mice would repair audibility deficits at threshold, and may also increase P1 amplitude. Whether it would repair the deficit at P4 is an interesting question, with neurobiological implications for understanding spatial acuity deficits in old mice. then switches to the other (the PP), the ES following from 1 ms to 300 ms later. An accelerometer under the cage detects the reflex flinch, its output integrated for 100 ms after the ES quantifies ASR amplitude. The data in Fig 4 show a minimal asymptotic PPI difference between the age groups at long ISI with a substantial early deficit in older mice extending out to at least 40 ms, even at 78 dB SPL. These data also suggest that the positive effect of increased carrier level continues to increase over longer intervals between the prepulse and the probe ES for all age groups. Separate angles are presented across days, whileISI, Carrier Level or Spectrum may be varied between or within days. Each stimulus condition is repeated 11 times. Trials are on average 20 seconds apart, the test session lasts about 50 minutes. Fig 1: PPI at 3 age groups, for separations of 22.5 to 180o across ISI at 60 dB SPL Fig 2: PPI at 45o, 10 ms ISI above, 50 ms below. Support from NIH (AG09524, DC05409) and the Schmitt Foundation for Integrative Brain Research JIson@bcs.rochester.edu