Download
1 / 11
120 likes | 413 Views
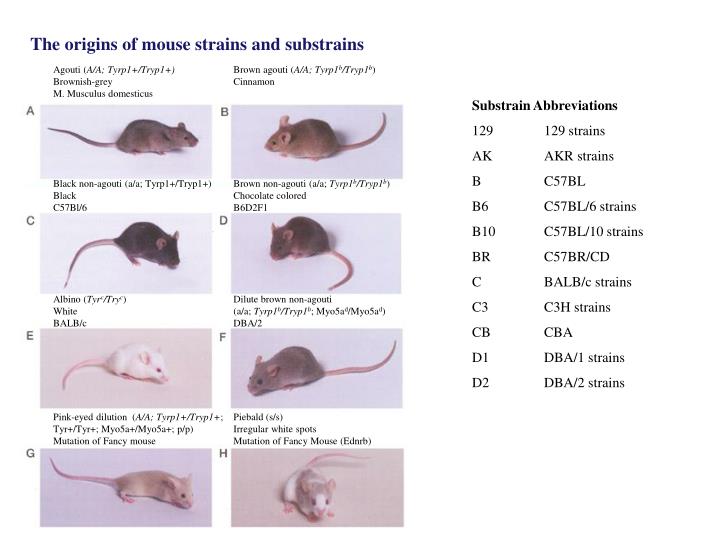
The origins of mouse strains and substrains. Agouti ( A/A; Tyrp1+/Tryp1+) Brownish-grey M. Musculus domesticus. Brown agouti ( A/A; Tyrp1 b /Tryp1 b ) Cinnamon. Substrain Abbreviations 129 129 strains AK AKR strains B C57BL B6 C57BL/6 strains B10 C57BL/10 strains BR C57BR/CD

E N D
The origins of mouse strains and substrains Agouti (A/A; Tyrp1+/Tryp1+) Brownish-grey M. Musculus domesticus Brown agouti (A/A; Tyrp1b/Tryp1b) Cinnamon Substrain Abbreviations 129 129 strains AK AKR strains B C57BL B6 C57BL/6 strains B10 C57BL/10 strains BR C57BR/CD C BALB/c strains C3 C3H strains CB CBA D1 DBA/1 strains D2 DBA/2 strains Black non-agouti (a/a; Tyrp1+/Tryp1+) Black C57Bl/6 Brown non-agouti (a/a; Tyrp1b/Tryp1b) Chocolate colored B6D2F1 Albino (Tyrc/Tryc) White BALB/c Dilute brown non-agouti (a/a; Tyrp1b/Tryp1b; Myo5ad/Myo5ad) DBA/2 Pink-eyed dilution (A/A; Tyrp1+/Tryp1+; Tyr+/Tyr+; Myo5a+/Myo5a+; p/p) Mutation of Fancy mouse Piebald (s/s) Irregular white spots Mutation of Fancy Mouse (Ednrb)
Generating new strains and substrains (i.e. inbred strains) Most research involving mice entails the use of inbred strains, not outbred stocks. The advantage of this is that it enables researchers in different parts of the world to work to reproduce experiments carried out in any country. By definition an inbred strain is generated by brother sister mating for 20 or more generations and the foundation stock from for the colony can be traced back to a single breeding pair. Once mice have been brother sister mated for 20 generations, then, on average, 98.6% of the genome should be homozygous. Therefore, all mice within a single inbred strain are both genetically identical (isogenic) and homozygous at virtually all loci. If the average mutation rate is taken as 5x 106, then there is a 90% probability that two substrains will differ at one or more loci after 16 generations of separation.
Embryo implantation in the mouse Following fertilization in the oviduct, the embryo undergoes several rounds of mitotic cell division, ultimately forming a ball of cells called a morula. At the late morula stage, the embryo enters the uterine lumen and transforms into a blastocyst that contains a cavity (called blastocoel) with two distinct cell populations, the inner cell mass (ICM) and the trophectoderm (the progenitor of trophoblast cells). Before implantation, the blastocyst escapes from its outer shell (the zona pellucida) and differentiates to produce additional cell types — the epiblast and the primitive endoderm. At this stage, the trophectoderm attaches to the uterine lining to initiate the process of implantation. E, embryonic day.
Isolation of Embryonic Stem (ES) cells In the blastocyst stage, cells of the inner cell mass (ICM) are totipotent, i.e. they can contribute to any embryonic tissue.However, they have lost the capacity to form extra-embryonic membranes. Blastocysts are isolated from an early mouse embryo. Cultured on feeder embryonic fibroblasts. Different from embryonic stem (ES) cells !! ES cell lines are derived in vitro to give pluripotent cells, i.e. not differentiated and can develop into all tissue types and cells of an embryo Each ES cell line must be tested for its ability to make chimeric mice. Blastocysts are fertilised, early developing embryos that have not yet attached to the mothers uterus The blastocyst on the left is c.a. 4 days after fertilization. Blastocyst attaches to the feeder layer and the ICM begins to grow Blastocyst Fibroblast feeder cells Expansion of the ICM to give ES cells Individual ES cell lines are tested for their ability to make chimeric mice
ES transfection/selection, blastocyst injection and germline transmission ES cell targeting:the process Homologous recombination refers to the exchange of DNA fragments between two DNA molecules at an identical site, which allows insertion of the transgene to be targeted to a specific location on the chromosome. Undifferentiated embryonic stem (ES) cells have the potential to differentiate into any type of cell within the developing organism. These cells are harvested from a blastocyst stage embryo and cultured in an in vitro environment. Transgene positive cells are identified and sorted using various selection techniques, including survival selection (positive-negative selection) and polymerase chain reaction (PCR) amplification. The modified ES cells are then injected directly into a normal blastocyst embryo.
Generation of a congenic line Cell line Strain source RW4 129X1/SvJ R1c 129X1/SvJ x J1 129S4/SvJae D3 129S2/SvPas AB1 129S7/SvEvBrd-Hprt TL1 129S6/SvEvTac TC1 129S6/SvEvTac W4 129S6/SvEv C1 129X1/SvJ E14TG2a 129P2/OlaHsd PluriStem BALB/C PluriStem C57BL/6N PluriStem DBA/2N
Genetic Nomenclature The Mouse Genome Database (MGD) serves as a central repository of gene names and symbols to avoid use of the same name for different genes or use of multiple names for the same gene. The Rat Genome Database serves the same purpose for that species. A key feature of mouse and rat nomenclature is the Laboratory Registration Code or Laboratory code, which is a code of usually three to four letters (first letter uppercase, followed by all lowercase), that identifies a particular institute, laboratory, or investigator that produced, and may hold stocks of, for example, a DNA marker, a mouse or rat strain, or were the creator of a new mutation. The gene/locus name serves as a unique identifier as it is the true unit of inheritance. It’s important because it identifies the gene as a member of a family (allowing additional information to be inferred), or not, and can be used to signify any gene orthologs, for example in human. A gene symbol must i) be unique, ii) be short (normally 3-5 characters), iii) begin with an uppercase letter (not a number), followed by all lowercase letters / numbers, iv) be italicized, v) comprise only Roman letters and Arabic numbers. By contrast, protein symbols use all uppercase letters and should not be italicized. Examples of Laboratory Codes J The Jackson Laboratory Mit Massachusetts Institute of Tech. Kyo Kyoto University Tac Taconics
Nomenclature for gene targeted loci Mutations that are the result of gene targeting by homologous recombination in ES cells are given the symbol of the targeted gene, with a superscript consisting of three parts: the symbol tm to denote a targeted mutation, a serial number from the laboratory of origin and the Laboratory code where the mutation was produced. Cftrtm1Unc the first targeted mutation of the cystic fibrosis transmembrane regulator (Cftr) gene produced at the University of North Carolina. Knock in mutations, in which all or part of the coding region of one gene is replaced by another, should be given a tm symbol and the particular details of the knock-in associated with the name in publications or databases. Where there has been a replacement of the complete coding region, the replacing gene symbol can be used parenthetically as part of the allele symbol of the replaced gene along with a Laboratory code and serial number. En1tm1(Otx2)Wrst the coding region of En1 was replaced by the Otx2 gene, originating from the W. Wurst laboratory. Gene trap mutations are symbolized in a similar way to targeted mutations. If the trapped gene is known, the symbol for the trapped allele will be similar to a targeted mutation of the same gene using the format Gt(vector content)#Labcode for the allele designation. Akap12Gt(ble-lacZ)15Brr a gene trap allele of the Akap12 gene, where the gene trap vector contains a phleomycin resistance gene (ble) and lacZ, the 15th analyzed in the laboratory of Jacqueline Barra (Brr). A transgene is any DNA that has been stably introduced into the germline of mice or rats. Such animals can be created can be by random insertion into the genome (usually by means of microinjection).
Conditional Gene Targeting: the Bacteriophage P1 System Conditonal gene targeting. Allows the gene to be removed at a chosen time or in a chosen tissue by regulating the expression of Cre-recombinase. Example A. The first exon of the gene is flanked by loxP sites, ‘floxed’. If this mouse is crossed with a transgenic mouse that expresses Cre early in development and in every tissue then pups will be born that have deleted the floxed gene in all tissues. Example B. If a liver specific Cre is used, such as an albumin promoter, then the floxed gene will only be deleted in liver. Alternatively, adenovirus that has the Cre cDNA can be injected into the tail vein of the mouse and It will localise to the liver where it removes the floxed allele.
Simplified nomenclature for conditional targeted animals Tg(Pdx1-Cre)1Agb a transgenic mouse line (Tg) carrying a Cre-transgene driven by Pdx1 promoter (Pdx1) This is the mouse line number 1 of this transgenic mouse kind (1) created by Anne Grapin-Botton (Agb) by insertion. Cftrtm1Unc For this example, lets pretend that the first targeted mutation of the cystic fibrosis transmembrane regulator (Cftr) gene was created by a conditional floxed construct Cftrtm1Unc :: Tg(Pdx1-Cre)1Agb This mouse carries both the Cre-transgene (under the expression of the Pdx1 promoter) and the conditional allele. Simplified nomenclature for the laboratory Cftr+/+ wild-type for both alleles Cftrflox/+ floxed on one allele and wild-type on the other Cftrdel/+ gene was deleted on one allele (by Cre; see below) and the other allele is wild-type Cftrdel/+:: Pdx1-Cre The Cre-recombinanse will delete the floxed allele in tissues were pdx1 is expressed