Download
1 / 34
340 likes | 353 Views
Learn how sequential gene expression determines segment identity in Drosophila embryos through regulatory genes. Discover the role of Polycomb and Trithorax complexes in maintaining cell identity and the impact of misexpressed Homeotic genes.

E N D
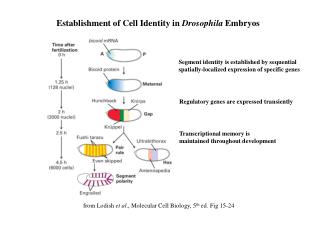
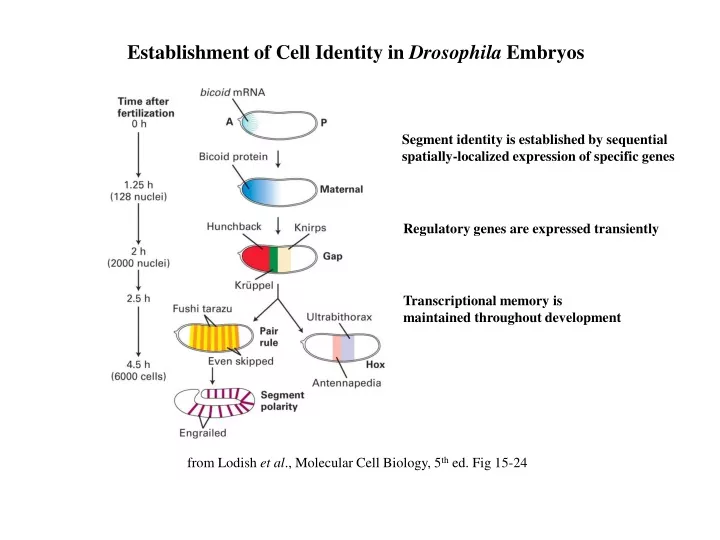
Establishment of Cell Identity in Drosophila Embryos Segment identity is established by sequential spatially-localized expression of specific genes Regulatory genes are expressed transiently Transcriptional memory is maintained throughout development from Lodish et al., Molecular Cell Biology, 5th ed. Fig 15-24
Misexpression of Homeotic Genes Lead to Morphological Abominations from Lodish et al., Molecular Cell Biology, 5th ed. Fig 15-25
Polycomb and Trithorax Complexes Prevents changes in cell identity by preserving transcription patterns Chromatin is altered in a heritable manner Components of PcG and trxG are commonly mutated in cancer Polycomb-group Proteins Maintains a silenced state Prevents chromatin remodelling Trithorax-group Proteins Maintains an active state Counteracts the action of PcG proteins Memory system composed of PcG and trxG complexes is linked to the histone code
Enzymatic Activities of Polycomb Group Complexes PcG proteins are recruited to Polycomb response elements EZH1 or EZH2 of PRC2 trimethylates H3K27 RING1A and RING1B of PRC1 ubiquitylates H2AK119 from Blackledge et al., Nature Rev.Mol.Cell Biol. 16, 643 (2015)
The Role of PcG Proteins in Chromatin Organization Forms PcG bodies Forms TADs Forms chromatin loops Cbx2 binds to H3K27me3 and condenses chromatin Modifies histones from Entrevan et al., Trends Cell Biol. 26, 511 (2016)
Targeting PRCs to Chromatin Two examples: PRC2 is recruited to inactive X chromosome by Xist PRC1 is targeted to unmethylated CpG islands and other mechanisms determine whether the association is stable from Blackledge et al., Nature Rev.Mol.Cell Biol. 16, 643 (2015)
Model for PRC-mediated Silencing from Blackledge et al., Nature Rev.Mol.Cell Biol. 16, 643 (2015) PRCs sample chromatin at regulatory elements and responds to the transcriptional state of the associated gene Transcription decreases residency or activity of PRCs Polycomb domains are established at silenced genes to limit stochastic reactivation
Propagation of Polycomb Domains on Newly Replicated Chromatin from Blackledge et al., Nature Rev.Mol.Cell Biol. 16, 643 (2015) EED binds H3K27me3 on old nucleosomes PRC2 trimethylates H3K27 on new nucleosomes
Formation of a Repressive Chromatin Hub PREs and promoters make contact and form chromatin loops CTCF and cohesin stabilize loops Chromatin loops are enriched in visible PcG bodies Loops could reinforce the memory of the silenced state from Bantignies and Cavalli, Trends Genet. 27, 454 (2011)
Segment-specific Localization of Genes in PcG Bodies PcG genes maintain the regional identity of segments by repressing Hox genes in specific regions Hox genes are organized in two clusters in Drosophila PcG bodies are subdomains of the nucleus that correlate with gene repression Antp and AbdB genes are silenced in the head (B) Antp and Ubx are silenced in the posterior (C) from Hodgson and Brock, Cell144, 170 (2011)
Epigenetics and Cancer Epigenetic dysregulation plays a causal role in cancer Epigenetic modifications are an efficient process to adapt to environmental and therapeutic pressures
Isocitrate Dehydrogenase Mutations Produce an Oncometabolite IDH converts isocitrate to a-ketoglutarate in the citric acid cycle Neomorphic IDH mutants produce R-2HG from a-ketoglutarate IDH1/2 mutants occur in certain types of cancer R-2HG inhibits a-ketoglutarate- dependent enzymes such as prolyl hydroxylases, lysine demethylases and TET IDH1/2 mutations lead to global DNA hypermethylation from Gagne et al, Trends Cell Biol., 27, 738 (2017)
Some Effects of IDH1/2 Mutations on Tumors Inhibits TET leading to hypermethylation Silencing of tumor suppressor genes Prevents CTCF binding Inhibits ATM phosphorylation cascade Decreased dsb repair Decreases endostatin production Stimulates HIF activity Promotes angiogenesis
IDH1/2 Mutations Activate Oncogenes from Grimmer and Costello, Nature529, 34 (2016) Mutant IDH1/2 produces R-2HG, which inhibits TET and leads to hypermethylation Reduced CTCF binding alters chromatin conformation A potent enhancer interacts with an oncogene promoter Modulating 3D chromatin structure may be widespread in cancer
HOTAIR Represses Genes in trans HOTAIR is a lncRNA expressed by the HOXC locus HOTAIR associates with PRC2 and LSD1 and recruits the complex to the HOXD locus HOTAIR acts in trans to repress the HOXD locus on a different chromosome from Kugel, Trends Biochem.Sci. 37, 144 (2012)
Trithorax-group Protein Mechanism of Action TrxG proteins maintain an active transcriptional state TrxG proteins promote H3K4me
Effect of Transcription on Epigenetic Modifications Transcription factor binding leads to histone modifications that promotes expression of neighboring genes from Furey and Sethupathy, Science 342, 705 (2013)
The Viable Yellow Agouti Locus Agouti promotes yellow pigment formation on black hair shaft Wild-type mice have brown fur due to Agouti expression from hair cell-specific promoter Avy contains an IAP insertion that contains a promoter expressed in all cells from Dolinoy, Nutr.Rev. 22(Suppl. 1),S7 (2008)
Avy is a Metastable Epiallele Avy can be modified in a variable and reversible manner Methylation status of IAP determines the activity of the ectopic promoter Ectopic Agouti expression causes yellow fur, obesity, diabetes and tumorigenesis Avy can be used as an epigenetic biosensor to study the nutritional and environmental influences on the fetal epigenome from Jirtle and Skinner, Nature Rev.Genet. 8, 253 (2007)
Maternal Nutrition Alters Gene Expression by Epigenetic Modification Feeding of pregnant Avy/a mice with methyl-rich supplements repress the ectopic Avy promoter Offspring of diet-supplemented mice have brown coat color and methylated IAP from Jirtle and Skinner, Nature Rev.Genet. 8, 253 (2007)
Epigenetic Inheritance in Mammals? There are several examples of nongenetic germline transmission of traits resulting from pesticides, metabolic state, and behavior The majority of sperm DNA is repackaged by protamines DNA is demethylated in the single-celled embryo DNA methylation is gradually reestablished in the early embryo A second round of DNA demethylation occurs in the developing germline from Hughes, Nature507, 22 (2014) The best evidence for epigenetic information transfer is through RNA
The Weismann Barrier Revisited Weismann – information flow can only flow from germ cells to somatic cells Information can be transferred from somatic cells to gern cells Can acquired characteristics be inherited? Transfer of EV contents to sperm from Chen et al., Nature Rev. Genet. 17, 733 (2016)
Intergenerational and Transgenerational Epigenetic Inheritance Exposures to metabolic state, stress, and pesticides have manifestations in subsequent generations Exposure during pregnancy affects somatic cells of F1 progeny and their developing germ cells which affects the F2 generation Exposure during adult life directly impacts the F1 progeny from Sales et al., Cell Metabolism 25, 559 (2017)
Non-genetic Influences to Progeny A fetus adapts to the environment to which it is exposed Undernutrition during gestation leads to T2D in offspring and in next generation Acquired behaviors can be transmitted across multiple generations
The Link Between Intrauterine Growth Retardation and T2D Infants born during the Dutch Hunger Winter had lower birth rates, but had impaired glucose tolerance later in life How much of this effect is epigenetic, and how much is due to parental control of eating and exercise habits?
Epigenetic Inheritance of Paternally-Acquired Traits DNA methylation is mostly erased, and TFs and histones are lost RNA mediates transgenerational effects through sperm Stress causes alterations in miRNA and piRNA following stress Sperm may acquire RNA via exosomes Offspring of zygotes injected with sperm RNA can recapitulate paternal phenotype Diet-induced changes in tRNA fragments account for nutrition- induced transgenerational effects from Chen et al., Nature Rev. Genet. 17, 733 (2016)
Parental Experiences Impact the Behavior of the Offspring from D’Urso and Brickner, Trends Genet. 30, 230 (2014) Stress reduces maternal care. Pups are more sensitive to stress and display reduced maternal care, even in the absence of stress The altered gene expression of target genes (GR in the hippocampus) is mediated by DNA methylation and histone modifications Expression patterns are inherited in future generations
Somatic Cell Reprogramming Pleuripotency genes in somatic cells have methylated CpG islands Epigenetic marks must be reset to generate induced pleuripotent stem (iPS) cells Repressive histone methylation marks must be removed, followed by removal of DNA methylation which activates the gene from Cedar and Bergman, Nature Rev.Genet. 10, 295 (2009)
Prion Epigenetics Prions template conformational conversion of other molecules of the same protein Prions are formed through an oligomeric nucleus, and the elongating polymer is severed by protein remodeling factors Prions are disseminated to daughter cells during cell division Good evidence for prion-mediated epigenetic inheritance in yeast from Halfmann and Lindquist, Science330, 629 (2010)
Stress Accelerates Prion Appearance Abrupt changes have consequences for protein folding Prion-free cells are adapted to environment 1, but poorly adapted to environment 2 Prion formation and disappearance provide fitness advantages in different environments from Halfmann and Lindquist, Science330, 629 (2010) Prions connect environmental conditions to acquisition and inheritance of new traits
The Metabolic State is Linked to Epigenetic Modifications Most chromatin-modifying enzymes use metabolites as co-factors The metabolic status of a cell can transduce a transcriptional response from Gut and Verdin, Nature502, 489 (2013)
The Complete Demethylation Pathway of 5mC 5mC is oxidated iteratively by TET 5hmC is reverted to unmodified C by passive dilution during DNA replication Oxidative products are excised by thymine DNA glycosylase and repaired by BER from Kohli and Zhang, Nature502, 472 (2013)
DNA Methylation Dynamics During Epigenetic Reprogramming Epigenetic memory must be erased for cells to achieve pleuripotency Maternal DNA undergoes passive demethylation in pre-implantation embryos Paternal genome is actively demethylated by TET3 DNA methylation patterns are re-established by de novo DNMTs at the blastocyst stage Demethylation also takes place during the embryonic phase of germline development Primordial germ cells are demethylated through a TET-independent and a TET-mediated oxidative pathway from Kohli and Zhang, Nature502, 472 (2013)
Incomplete Erasure of DNA Methylation Repetitive elements Some promoters Short-term transgenerational epigenetic effects can me mediated by RNA