Download
1 / 29
370 likes | 1.55k Views
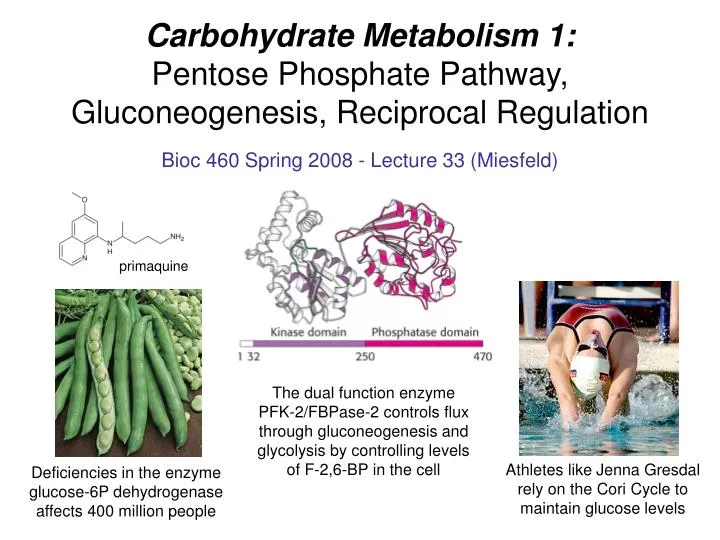
Carbohydrate Metabolism 1: Pentose Phosphate Pathway, Gluconeogenesis, Reciprocal Regulation. Bioc 460 Spring 2008 - Lecture 33 (Miesfeld). primaquine. The dual function enzyme PFK-2/FBPase-2 controls flux through gluconeogenesis and glycolysis by controlling levels of F-2,6-BP in the cell.

E N D
Carbohydrate Metabolism 1:Pentose Phosphate Pathway, Gluconeogenesis, Reciprocal Regulation Bioc 460 Spring 2008 - Lecture 33 (Miesfeld) primaquine The dual function enzyme PFK-2/FBPase-2 controls flux through gluconeogenesis and glycolysis by controlling levels of F-2,6-BP in the cell Athletes like Jenna Gresdal rely on the Cori Cycle to maintain glucose levels Deficiencies in the enzyme glucose-6P dehydrogenase affects 400 million people
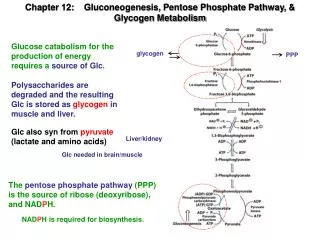
Key Concepts: The Pentose Phosphate Pathway • The pentose phosphate pathway takes place entirely within the cytoplasm and is also known as the hexose monophosphate shunt or phosphogluconate pathway. • The most important function of the pentose phosphate pathway is to reduce two molecules of NADP+ to NADPH (nicotinamide adenine dinucleotide phosphate) for each glucose-6-phosphate that is oxidatively decarboxylated to ribulose-5-phosphate. • NADPH is functionally similar to NAD+ however, NADPH is the primary reductant in the cell, whereas, NAD+ is the primer oxidant. NADPH is critical to maintaining reduced glutathione levels in cells which is required to minimized damage from reactive oxygen species. • The pentose phosphate pathway is also responsible for producing ribose-5-phosphate which provides the ribose sugar backbone that anchors the nucleotide base to DNA and RNA polymers.
We will cover three primary pathways related to carbohydrate metabolism in non-photosynthetic organisms: • Pentose phosphate pathway • Gluconeogenesis • Glycogen metabolism Metabolism of ribose sugars in the pentose phosphate pathway is used to generate NADPH and to provide the carbohydrate component of nucleotides. The major sources of carbon in gluconeogenesis are amino acids and glycerol in animals, and glyceraldehyde-3-phosphate (GAP) in plants.
Pathway Questions 1. What does the pentose phosphate pathway accomplish for the cell? • The oxidative phase generates NADPH which is required for many biosynthetic pathways and for detoxification of reactive oxygen species. • The nonoxidative phase interconverts C3, C4, C5, C6 and C7 monosaccharides to produce ribose-5P for nucleotide synthesis, and also to regenerate glucose-6P to maintain NADPH production by the oxidative phase. 2. What is the overall net reaction of the pentose phosphate pathway when it is utilized to generate the maximum amount of NADPH? 6 Glucose-6P + 12 NADP+ + 12 H2O → 5 Glucose-6P + 12 NADPH + 12 H+ + 6 CO2
Pathway Questions 3. What are the key enzymes in the pentose phosphate pathway? Glucose-6P dehydrogenase (G6PD)– enzyme catalyzing the first reaction in the pathway which converts glucose-6P to 6-phosphogluconolactone. This reaction is the commitment step in the pathway and is feedback-inhibited by NADPH. Defects in glucose-6P dehydrogenase cause a dietary condition called favism. Transketolase and Transaldolase - together these two enzyme catalyze the reversible "carbon shuffle" reactions of the nonoxidative phase of the pathway. These are the same enzymes used in the Calvin Cycle to regenerate ribulose-5P from glyceraldehyde-3P.
Pathway Questions 4. What are examples of the pentose phosphate pathway in real life? Glucose-6P dehydrogenase deficiency is the most common enzyme deficiency in the world and affects over 400 million people. A 90% decrease in enzyme activity results in the inability of red blood cells to produce enough NADPH to protect the cells from reactive oxygen species that are generated by anti-malarial drugs (primaquine) and by compounds in fava beans (vicine). Malaria-infected Anopheles mosquito biting a human A big bowl of fava beans
Two Phases of the Pentose Phosphate Pathway The pentose phosphate pathway can be divided into two phases, the oxidative phase, which generates NADPH, and the nonoxidative phase, which interconverts C3, C4, C5, C6 and C7 sugar phosphates using many of the same "carbon shuffle" reactions we saw in the Calvin cycle.
Three enzymatic reactions in the oxidative phase 1. Oxidation of glucose-6P by the enzyme glucose-6P dehydrogenase (G6PD) to 6-phosphogluconolactone is coupled to the reduction of NADP+ resulting in the formation of one molecule of NADPH. This is the commitment step in the pathway. 2. 6-phosphogluconolactone is hydrolyzed by lactonase to produce the open chain monosaccharide 6-phosphogluconate. 3. 6-phosphogluconate is then oxidized and decarboxylated by 6-phosphogluconate dehydrogenase to generate ribulose-5P, CO2 and the second molecule of NADPH. 1 2 3
1 2 3 4 6 5 1 2 3 4 5 The non-oxidative phase of the PPP In cells that require high levels of NADPH for biosynthetic reactions, the ribulose-5P produced in the oxidative phase needs to be converted back into glucose-6P to maintain flux through the glucose-6P dehydrogenase reaction. The carbon shuffle reactions of the nonoxidative phase are used to regenerate glucose-6P using the same transketolase and transaldolase enzyme reactions as the Calvin Cycle. Where did the 6 carbons go?
Metabolic flux through the Pentose Phosphate Pathway is tightly-regulated 1. If increased NADPH is required for biosynthetic pathways, or to provide reducing power for detoxification, then fructose-6P and glyceraldehyde-3P are used to resynthesize glucose-6P and thereby maintain flux through the oxidative phase of the pathway. 2. If cells need to replenish nucleotide pools due to high rates of DNA and RNA synthesis, then the bulk of ribulose-5P is converted to ribose-5P to stimulate nucleotide biosynthesis. 3. If ATP levels in the cell are low, and NADPH levels are not limiting, then glucose-6P dehydrogenase is inhibited and the pentose phosphate pathway is bypassed so that glucose-6P can be metabolized directly by the glycolytic pathway.
Regulation of the G6PD activity controls flux through the glycolytic pathway and pentose phosphate pathways When the rates of NADPH-dependent biosynthetic reactions are high in the cytosol, then the [NADP+]/[NADPH] ratio increases, leading to allosteric activation of glucose-6P dehydrogenase activity by NADP+ which increases flux through the pentose phosphate pathway. Increased levels of NADPH compete with NADP+ for binding to glucose-6P dehydrogenase, thereby reducing the activity of the enzyme. This results in decreased flux through the pentose phosphate pathway and the available glucose-6P is then metabolized by the glycolytic pathway to increase production of ATP.
Glucose-6P dehydrogenase deficiency in humans The pentose phosphate pathway is responsible for maintaining high levels of NADPH in red blood cells (erythrocytes) for use as a reductant in the glutathione reductase reaction. Glutathione is a tripeptide that has a free sulfhydryl group which functions as an electron donor in a variety of coupled redox reactions in the cell. Glutathione reductase uses two electrons from NADPH to maintain glutathione in the reduced state (GSSG → 2 GSH).
Glucose-6P dehydrogenase deficiency in humans In erythrocytes, electrons from glutathione are used to reduce harmful reactive oxygen species and hydroxyl free radicals. When erythrocytes are exposed to chemicals that generate high levels of superoxide radicals, GSH is required to reduce these damaging compounds. The pentose phosphate pathway in erythrocytes normally provides sufficient levels of NADPH to maintain the GSH:GSSG ratio at about 500:1.
Glucose-6P dehydrogenase deficiency in humans Result of observations made 30 years earlier – the anti-malarial drug primaquine induced acute hemolytic anemia (red blood cell lysis) in a small percentage of people who had been given primaquine prophylatically. Primaquine inhibits growth of the malarial parasite in red blood cells by creating a hostile environment (reactive oxygen species). The biochemical basis for this drug-induced illness was found to be lower than normal levels of NADPH due to a G6PD deficiency. The acute hemolytic anemia seen in individuals with G6PD who are treated with primaquine explains the symptoms of favism. One of the compounds in fava beans is vicine, a toxic glycoside that induces oxidative stress in erythrocytes. What might explain the observation that cultures with high amounts of fava beans in the diet were associated (in ancient times) with low malaria rates?
Key Concepts: Gluconeogenesis • The importance of gluconeogenesis is to provide glucose for cells from non-carbohydrate precursors, primarily the carbon backbone of amino acids, plants use gluconeogenesis to convert GAP to glucose. • Three steps in glycolysis must be bypassed by gluconeogenic enzymes in order to overcome large G differences. Two of the steps simply reverse the reaction (fructose-1,6-bisphosphatase and glucose-6-phosphatase), whereas, another step requires two bypass enzymes (pyruvate carboxylase, PEP carboxykinase). • Flux through gluconeogenesis and glycolysis is reciprocally-regulated to prevent futile cycling (burning up ATP). Reciprocal regulation at the PFK-1 (glycolysis) and F-1,6-BPase (gluconeogenesis) is controlled by the allosteric regulator F-2,6-bisphosphate, as well as, energy charge (ATP/AMP), and citrate levels. • The Cori Cycle recycles lactate produced in anaerobic muscle cells during exercise by exporting it to the liver where it is converted to pyruvate and used to synthesize glucose by gluconeogenesis.
Pathway Questions 1. What does gluconeogenesis accomplish for the organism? The liver and kidney generate glucose from noncarbohydrate sources (lactate, amino acids, glycerol) for export to other tissues that depend on glucose for energy, primarily the brain and erythrocytes. Plants use the gluconeogenic pathway to convert GAP, the product of the Calvin Cycle, into glucose which is used to make sucrose and starch.
Pathway Questions 2. What is the overall net reaction of gluconeogenesis? 2 pyruvate + 2NADH + 4ATP + 2GTP + 6H2O → Glucose + 2NAD+ + 2H+ + 4ADP + 2GDP + 6Pi 3. What are the key enzymes in gluconeogenesis? Pyruvate carboxylase is a mitochondrial enzyme that catalyzes a carboxylation reaction converting pyruvate to oxaloacetate.Phosphoenolpyruvate carboxykinase (PEPCK)converts oxaloacetate to phosphoenolpyruvate (PEP) using the energy released by decarboxylation and GTP hydrolysis. Transcription of the PEPCK gene is regulated by hormones.Fructose-1,6-bisphosphatase-1 (FBPase-1) catalyzes the dephosphorylation of fructose-1,6BP to form fructose-6P; this is the bypass reaction for PFK-1 in glycolysis. Glucose-6-phosphatase is an enzyme in liver and kidney cells (not present in muscle cells) that catalyzes the dephosphorylation of glucose-6P to form glucose which can be exported out of the cell.
Pathway Questions Application of gluconeogenesis in real life Monitoring blood glucose levels throughout the day is critical to diabetics who need insulin injections. Glucose monitoring devices are based on an assay using the enzyme glucose oxidase which produces gluconate and hydrogen peroxide (H2O2) from glucose. The level of H2O2 in the sample is detected by an indicator dye that is oxidized in a reaction catalyzed by peroxidase.
Glycolysis and gluconeogenesis are opposing pathways that serve the critical function of degrading or synthesizing glucose in response to energy demands in the cell. These two pathways share seven of the same enzymes, with additional pathway-specific enzymes required at the three key regulatory steps. Two of the bypass enzymes in gluconeogenesis,fructose-1,6-bisphosphatase-1 (FBPase-1) and glucose-6-phosphatase, simply reverse the reaction However, 4 extraATP/GTP, and pyruvate carboxylase and phosphoenolypyruvate carboxykinase (PEPCK), are required to catalyze the bypass reaction that converts pyruvate to PEP.
Pyruvate carboxylase is a mitochondrial enzyme that requires the cofactor biotin to function as a carboxyl group carrier in a two step enzyme reaction. Pyruvate carboxylase is activated by acetyl CoA and has an important role in supplying OAA to the citrate cycle when acetyl CoA levels are high and the energy charge in the cell is low. The cellular location of PEPCK differs depending on the species. Humans contain two distinct PEPCK genes that encode mitochondrial and cytosolic PEPCK enzymes.
Reciprocal regulation of PFK-1 and FBPase-1 The activities of PFK-1 and FBPase-1 are regulated by the allosteric effectors AMP, citrate and fructose-2,6-bisphosphate (F-2,6-BP), but in a reciprocal manner. Reciprocal regulation refers to the fact that the same regulatory molecule has opposite effects on two enzymes that control a shared step in two reaction pathways. For example, when energy charge in the cell is low, AMP levels are high leading to activation of PFK-1 (increased flux through glycolysis) and inhibition of FBPase-1 (decreased flux through gluconeogenesis). This makes sense because the pyruvate generated by glycolysis can then be used in the energy conversion pathways to replenish ATP, while at the same time, glucose synthesis is shutdown resulting in a build-up of pyruvate. What is the metabolic logic of reciprocal regulation by citrate?
Reciprocal regulation of PFK-1 and FBPase-1 The allosteric regulator F-2,6-BP is an even more potent regulator of these two enzymes than either AMP or citrate. F-2,6-BP not a metabolic intermediate in either the glycolytic or gluconeogenic pathways, instead it is an allosteric regulator that activates PFK-1 and inhibits FBPase-1. In the presence of F-2,6-BP, the affinity of PFK-1 for its substrate fructose-6P is 25 times higher than it is in the absence of F2,6BP. Looking at the activity curves for FBPase-1 in the presence and absence of F-2,6-BP it can be seen that the affinity of FBPase-1 for its substrate fructose-1,6BP is 15 times lower in the presence of F-2,6-BP.
Levels of F-2,6-BP in the cell are controlled by a dual function enzyme called PFK-2/FBPase-2 The amount of F-2,6-BP in the cell is regulated by hormone signaling through glucagon and insulin which control the activity of a dual function enzyme containing two catalytic activities, 1) a kinase activity called phosphofructokinase-2 (PFK-2) that phosphorylates fructose-6P to form F-2,6-BP, and 2) a phosphatase activity called fructose-2,6-bisphosphatase (FBPase-2) that dephosphorylates F-2,6-BP to form fructose-6P.
Levels of F-2,6-BP in the cell are controlled by a dual function enzyme called PFK-2/FBPase-2 When the PFK-2/FBPase-2 dual function enzyme is unphosphorylated, then the PFK-2 activity in the enzyme is stimulatedand the FBPase-2 activity is inhibited, resulting in the net phosphorylation of fructose-6P to produce more F-2,6-BP which stimulates glycolytic flux. In contrast, when PFK-2/FBPase-2 is phosphorylated, the activity of PFK-2 is inhibited and the activity of FBPase-2 is stimulated.
Levels of F-2,6-BP in the cell are controlled by a dual function enzyme called PFK-2/FBPase-2 Activation of the glucagon receptor in liver cells results in stimulation of protein kinase A signaling which leads to phosphorylation of the PFK-2/FBPase-2 enzyme, thereby leading to decreased levels of F-2,6-BP and increased activity of the gluconeogenic enzyme FBPase-1.
Levels of F-2,6-BP in the cell are controlled by a dual function enzyme called PFK-2/FBPase-2 In contrast, insulin signaling stimulates protein phosphatase-1 activity resulting in the dephosphorylation of the PFK-2/FBPase-2 enzyme leading to higher levels of F-2,6-BP and activation of the glycolytic enzyme PFK-1.
The Cori Cycle The Cori cycle provides a mechanism to convert lactate produced by anaerobic glycolysis in muscle cells to glucose using the gluconeogenic pathway in liver cells. Although it costs four high energy phosphate bonds to run the Cori cycle (the difference between 2 ATP produced by anaerobic glycolysis and 4 ATP and 2 GTP consumed by gluconeogenesis), the benefit to the organism is that glycogen stores in the muscle can be quickly replenished following prolonged exercise. .
The Cori Cycle is important for peak performance Studies on athletes have shown that within 30 minutes of completing a vigorous workout, the majority of lactate produced during anaerobic glycolysis in the muscle has been converted to glucose in the liver and used to replenish muscle glycogen stores. In fact, the reason you should "warm down" after exercise (same movement but under aerobic conditions) is to enhance circulation so that lactate will be cleared from the muscle and be used in the liver for glucose synthesis via the Cori cycle.