Download
1 / 46
460 likes | 712 Views
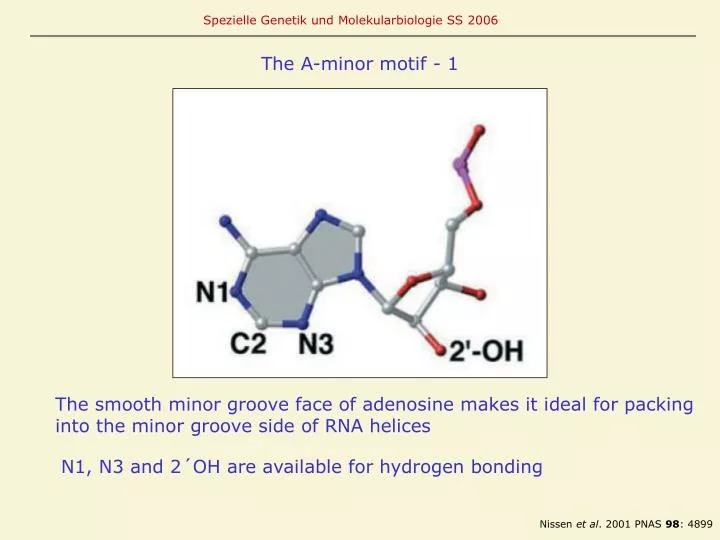
The A-minor motif - 1. The smooth minor groove face of adenosine makes it ideal for packing into the minor groove side of RNA helices. N1, N3 and 2´OH are available for hydrogen bonding. Nissen et al . 2001 PNAS 98 : 4899. N3 "inside" and 2´OH "outside".

E N D
The A-minor motif - 1 The smooth minor groove face of adenosine makes it ideal for packing into the minor groove side of RNA helices N1, N3 and 2´OH are available for hydrogen bonding Nissen et al. 2001 PNAS 98: 4899
N3 "inside" and 2´OH "outside" The A-minor motif – adenosine specific interactions N3 and 2´OH "inside" (For A-U no H-bond with N3) Nissen et al. 2001 PNAS 98: 4899
The A-minor motif – adenosine preferred interactions N3 and 2´OH "outside Nissen et al. 2001 PNAS 98: 4899
The decoding problem Ogle et al. 2003 TIBS 28: 259
A two step recognition mode Ogle & Ramakrishnan 2005 Ann. Rev. Biochem 74: 129
Phe-tRNAPhe (Prf 16/17) (fluorescent tRNA derivative) 1. Remove Y (wybutine) by acid treatment 2. React remaining aldehyde group at sugar with proflavine Gromadski & Rodnina 2004 Mol. Cell 13: 191
mant-GTP (fluorescent GTP derivative) 3´-O-(N-methylanthraniloyl)-2-deoxyguanosine triphosphate (structure for 2´/3´-O-(N-methylanthraniloyl)-[-thio] guanosine triphosphate, triethyl ammonium salt) Gromadski & Rodnina 2004 Mol. Cell 13: 191
Stopped flow - Under pneumatic drive activation, the two small volumes of solutions are driven from high performance syringes through a high efficiency mixer. - The resultant mixture passes through a measurement flow cell and into a stopping syringe. - Just prior to stopping, a steady state flow is achieved. - As the solution fills the stopping syringe, the plunger hits a block, causing the flow to be stopped instantaneously. - Using appropriate techniques, the kinetics of the reaction can be measured in the cell.
poly(U) programmed poly(A) programmed Example 1 Binding of EF-Tu-GTP-Phe-tRNAPhe (Prf16/17) to 70S ribosomes Subsequent fitting using a reasonable model for the reaction yields kinetic parameters Rodnina et al. 1994 Biochemistry 33: 12267
Example 2 Binding of EF-Tu-mantGTP-Phe-tRNAPhe to 70S ribosomes 1: poly(U) programmed with Ac-Phe-tRNAPhe in P-site 2: poly(A) programmed with tRNALys in P-site Rodnina et al. 1995 EMBO J 14: 2613
Measurement of peptide bond formation [3H]fMet-tRNA + ternary complex of [14C]Phe-tRNA rapid quenching with 0.8 M KOH (similar to stopped flow) determination of dipeptide formed Gromadski & Rodnina 2004 Mol. Cell 13: 191
Kinetics constants for inital selection Gromadski & Rodnina 2004 Mol. Cell 13: 191
Initial selection vs. proofreading Gromadski & Rodnina 2004 Mol. Cell 13: 191
Conformational changes of EF-Tu upon GTP hydrolysis EF-Tu-GDP EF-Tu-GTP Molecular Biology of the Gene
The tRNA binding site is only present in EF-Tu-GTP Molecular Biology of the Gene
Ovierview of structure ASL: gold, U6 "mRNA": purple, helix 44: cyan, 530 loop: light green, helix 34: light blue, S12: tan, P-site tRNA mimic (helix 6 from neighbouring molecule): dark blue, P-site mRNA mimic (3´ end of 16S rRNA): dark blue 3´AAG 5´ anticodon Ogle et al. 2001 Science 292: 899
Changes upon binding of cognate tRNA A1492 and A1493 flip out of internal loop of helix 44 G530 goes from syn to anti conformation Ogle et al. 2001 Science 292: 899
Interactions of A1493 at the first position class I A-minor interaction, requires Watson-Crick base bair Ogle et al. 2001 Science 292: 899
Interactions of A1492 and G530 at the second position class II A-minor interaction of A1492 interaction network requires Watson-Crick base pair Ogle et al. 2001 Science 292: 899
Interactions of G530 at the third position Mg2+ No Watson-Crick base pair required for interaction Ogle et al. 2001 Science 292: 899
Paromomycin induces similar changes As streptomycin, paromomycin reduces translational fidelity, but binding site is different Ogle et al. 2001 Science 292: 899
Near-cognate ASLs do not yield defined electron density anticodon: 3´GAG 5´ no binding? disordered binding? Ogle et al. 2002 Cell 111: 721
Electron density is obtained with paromomycin A1492 and A1493 flipped out G530 in anti conformation Ogle et al. 2002 Cell 111: 721
Conformation changes of 30S indicate binding of near-cognate ASL Ogle et al. 2002 Cell 111: 721
Conformation change are different in the presence of paromomycin anticodon: 3´GAG 5´: first & third position wobble Ogle et al. 2002 Cell 111: 721
Similar differences are observed for another near-cognate ASL anticodon: 3´AGG 5´: second & third position wobble Ogle et al. 2002 Cell 111: 721
Cognate tRNA induces closing also without paromomycin anticodon 3´AAG 5´: third position wobble Ogle et al. 2002 Cell 111: 721
Binding of first position wobble pair (with paromomycin) lack of complementary surface (van der Waals interactions) dehydration of 2´OHs required (no space for water) no compensation Ogle et al. 2002 Cell 111: 721
Binding of second position wobble pair (with paromomycin) gray/black indicates regular conformation of G-U wobble pair Ogle et al. 2002 Cell 111: 721
No well defined density for either syn or anti conformation of G530 Ogle et al. 2002 Cell 111: 721
Binding affinities competition experiments with 70S ribosomes Paromomycin increases the affinity of cognate ASL but not of near-cognate ASLs! Ogle et al. 2002 Cell 111: 721
Reasons? Cognate ASL always induces domain closure. Sufficient energy gain to pay the energy costs for domain closure. Paromomycin should lower those costs, since e.g. the entropic costs for fixing A1492/1493 are already payed by paromomycin binding. Near-cognate ASL binding alone does not gain enough energy to pay cost for domain closure. Only if part of the costs is payed by parmomycin, domain closure is possible. The total gain of energy seems to be similar with and without paromomycin for near-cognate ASL binding. Ogle et al. 2002 Cell 111: 721
Energy penalties for wobble base pairs Ogle et al. 2002 Cell 111: 721
The hybrid model Moazed & Noller 1989 Nature 342: 142
Cryo-EM of EF-Tu-tRNA complexes with the ribosome kirromycin stalled EF-Tu-GDP-tRNA complex side view top view mostly w/o EF-Tu mostly with EF-Tu Valle et al. 2002 EMBO J 21: 3557
Kirromycin stalled complexes are slightly different Valle et al. 2002 EMBO J 21: 3557
Ternary complex crystal structure does not fit density of it at ribosome Valle et al. 2002 EMBO J 21: 3557
tRNA needs to be distorted fitting of tRNA from EF-Tu-GTP-tRNA complex fitting of A-site tRNA in 70S ribosome complex fitting of a chimera of both structures Valle et al. 2002 EMBO J 21: 3557
Accomodation involves relaxation of tRNA Faster for cognate tRNA due to restricted conformational space? Ogle & Ramakrishnan 2005 Ann. Rev. Biochem 74: 129
EF-Tu GTPase activation? tRNA? sarcin-ricin-loop (SRL)? Ogle & Ramakrishnan 2005 Ann. Rev. Biochem 74: 129
Conditional streptomycin dependent (CSD) mutants CSD mutants grow well in rich medium CSD mutants require streptomycin for growth in minimal medium Gorini & Kataja 1964 PNAS 51: 487
Phenotypes of CSD mutants mutation maps in region of OTC gene Gorini & Kataja 1964 PNAS 51: 487
Streptomycin induces mistranslation in vitro ribosomes + 20 amino acids, one labeled in each experiment Ile: AUY, AUA Leu: CUN, UUR Ser: UCN, AGY Davies et al. 1964 PNAS 51: 883
restrictive and ram mutations Restrictive mutations abolish read-through at leaky mutations. Map in rpsL gene (protein S12) required for streptomycin resistance. Restrictive mutation have a hyperaccurate phenotype in translation. ram mutations (ribosomal ambiguity) revert restrictive phenotypes. Second site mutations that change proteins S4 or S5. ram single mutants have increased read-through at nonsense mutations. Ogle & Ramakrishnan 2005 Ann. Rev. Biochem. 74: 129
restrictive and ram mutations may influence closure movement Ogle & Ramakrishnan 2005 Ann. Rev. Biochem. 74: 129
The Hirsh suppressor (tRNATrp G24A) may increase flexibility Hirsh suppressor allows read-through at UGA with Trp Ogle & Ramakrishnan 2005 Ann. Rev. Biochem. 74: 129






















