Download

1 / 38
380 likes | 399 Views
Discover the detailed insights from the Aspergillus genomes, including karyotyping details, synteny maps, and gene expression patterns in response to environmental factors. Explore unique genes, ortholog clusters, and more.

E N D
What The Aspergillus Genomes Have Told Us William C. Nierman The Institute for Genomic Research Rockville, MD
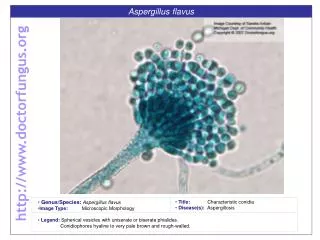
Electrophoretic Karyotyping 5 day run Af Sp Sc 5.7 5.0 4.6 1x 4.0 3.5 3.5 1.8 CHEF DRII 1.2% CGA, 1x TAE, 14C, 1.8 V/cm: 2200 s, 48 h; 2200-1800 s, 68 h sizes in Mb
Aspergillus fumigatus karyotype 1,789 Kb 3,779 Kb 2,021 Kb 3,992 Kb 4,018 Kb 4,834 Kb 4,891 Kb 3,933* Kb
Optical Analysis • Molecule maps generated from images of single DNA molecule digested with NheI • Resolution (avg fragment size) 8.28kb • Total coverage: 8,987 Mbase, or 300x • Total of 8 chromosomes • Total size: 29.189 Megabases
Aspergillus fumigatusChromosomes 1 4.9 Mb 2.7 2.2 2 4.8 Mb 1.8 3.0 3 1.3 2.8 4.0 Mb rRNA 4 0.7 2.5 0.4 3.9 Mb 0.3 5 1.2 3.9 Mb 2.6 6 1.3 3.6 Mb 2.5 Mitochondrion 7 1.3 2.0 Mb 0.7 32 Kb 8 0.8 1.0 1.8 Mb Presumed centromeric area Telomere
TIGR Autoannotation vs Sanger Curated Annotation • Status Count • Total Sanger genes analyzed 360 • Same gene structure 137 • Different gene structure 177 • Sanger missing in TIGR annotation 37 • Sanger matches multiple TIGR annotations 2 • Sanger, TIGR annotations opposite strands 7 • TIGR missing in Sanger annotation 12 • TIGR matches multiple Sanger annotations 9
Using Ortholog Clusters to Identify Potential Annotation Problems
Using Ortholog Clusters to Identify Potential Annotation Problems Different exon number due to annotation discrepancy
In some cases, differences in exon number are real We need to be able to distinguish annotation inconsistencies from real, interesting phenomena
Expression profiling analysis to study • Pathogenesis • Response to fungicidal drugs • Temperature-dependent gene expression - A. fumigatus is an environmental species can grow at temperatures as high as 55ºC can survive at temperatures up to 70ºC. - It is commonly isolated from metabolically heated compost heaps
The Beast: Microarray Robot from Intelligent Automation <http://www.ias.com>
Scanning Hybridization Reference sample Query sample Multi-experiment comparison Obtain signal intensity values from images Data Normalization and analysis Microarray data analysis Software freely available at,< http://www.tigr.org/software >
Temperature shift experiments • Two shift experiments • 30ºC to 37ºC shift • 30ºC to 48ºC shift • Design • A. fumigatus was grown in a rich medium at 30ºC for two days from conidia, and shifted to 37ºC or to 48ºC. • Samples were taken throughout a time course. • Samples were prepared in Greg May’s Lab
A number of genes of various functional roles express differentially at each temperature. • More genes are shifted to down-regulation than up-regulation at 48˚C in comparison to 30˚C. • More genes are turned up at 37˚C when temperature was shifted from 30˚C. This suggests that the fungus has more variety of activities at 37˚C than does at the other temperatures, and it is least active at 48˚C.
heat shock proteins 30ºC to 37ºC784 genes
heat shock proteins 30ºC to 48ºC257 genes
More heat shock and stress-responsive genes (ex. those coding for heat shock proteins and chaperons) are highly expressed at 48˚C than are at lower temperatures, indicating that the fungus is under heat stress. • More putative virulence genes (ex. those coding for the proteins responsive to oxidative stress and host immune system and for toxin production) are highly expressed at 37˚C, although there is no contact with host cells. While predicted function from each gene should be experimentally verified, we suggest from this study that temperature is a key environmental signal for the organism that triggers gene regulation cascades that may ultimately lead to adaptation to a specific new environment.
Many transposases, especially those of Mariner-4 type, are highly expressed at 48˚C. It will be interesting to see if the high expression of the transposases actually leads to the transposition events of the transposons.
Transposons in A. fumigatus Dispersed in the genome
Overview – comparative statistics The ortholog was computed by performing an all vs. all BlastP of the three proteomes with a cut-off of 1 x e-15 (no length requirement). The mutual best hits were then organized into clusters based on shared protein nodes.
Aspergillus fumigatus Unique Genes • Vast majority are hypothetical • Includes • Several transcriptional regulators • A chaperonin • An hsp 70 related protein • ArsC, arsenate reductase • Teichoic Acid Biosynthetic Protein
Comparative Genomic Hybridization (CGH) • Competitive hybridization between two genomic DNA • Uses microarray to score the presence of genes relative to the reference on the microarray • Provides a quick and easy way of comparing the gene content of a reference organism relative to an unsequenced CLOSE relative
A. fumigatus vs. A. fischerianus • Within same cluster by large subunit rRNA analysis • Average DNA identity of ~ 90% based on 4X contigs of A. fischerianus • A. fischerianus rarely identified as a pathogen • A. fischerianus possesses a known sexual cycle
A. fumigatus vs. A. fischerianus • Relative to A. fumigatus, A. fischerianus is missing 700 genes • 13 Secondary metabolite genes • 28 Transcription regulators and protein kinases • 21 Transporters • 199 Metabolic and other proteins • 400 Hypothetical proteins
A. fumigatus vs. A. fischerianusSecondary Metabolite Gene Summary • Relative to A. fumigatus, A. fischerianus is missing • 3 of 7 DMAT genes • 6 of 14 PKS genes • 1 of 15 NRPS genes
Additional related genomic projects underway or soon to be initiated • Comparative analysis of Aspergillus fumigatus AF293 and CEA10 • Sequencing of Aspergillus flavus • Sequencing of Aspergillus terreus • Sequencing of Aspergillus clavatus • Sequencing of Aspergillus fischerianus • CGH of Neosartorya fennelliae with A. fumigatus • CGH across multiple A. fumigatus strains
Aspergillus fumigatus AF293 Joan Bennett Matt Berriman Jean Paul Latge Paul Dyer Paul Bowyer Neil Hall David Denning Michael Anderson Arnab Pain Goeff Robson Javier Arroyo Goeff Turner David Archer Aspergillus nidulans – James Galagan Aspergillus oryzae – Masayuki Machida
TIGR Sequencing and Closure Tamara Feldblyum Hoda Khouri Microarray H. Stanley Kim Dan Chen Annotation Jennifer Wortman Jiaqi Huang Resham Kulkarni Natalie Fedrova Claire Fraser NIAID and Dennis Dixon