Download
1 / 29
290 likes | 459 Views
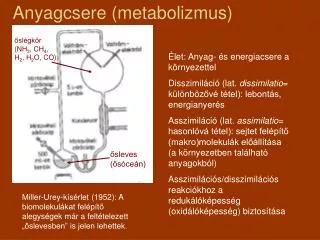
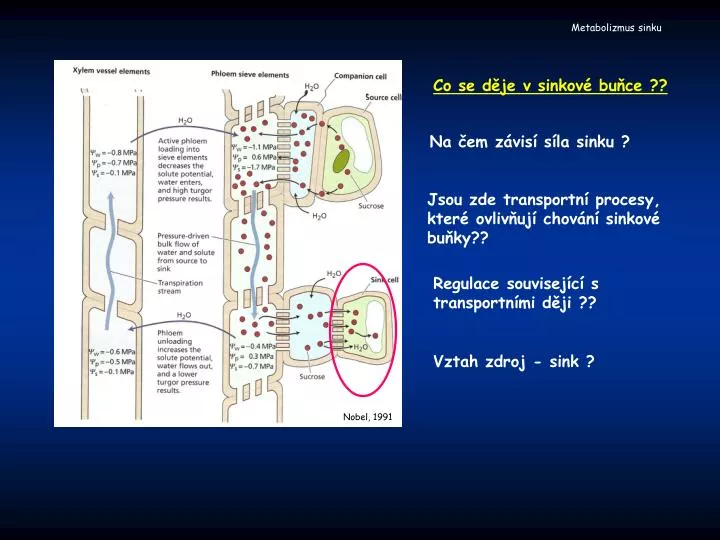
Nobel, 1991. Metabolizmus sinku. Co se děje v sinkové buňce ??. Na čem závisí síla sinku ?. Jsou zde transportní procesy, které ovlivňují chování sinkové buňky??. Regulace související s transportními ději ??. Vztah zdroj - sink ?. Metabolizmus sinku. Charakteristika sinku.

E N D
Nobel, 1991 Metabolizmus sinku Co se děje v sinkové buňce ?? Na čem závisí síla sinku ? Jsou zde transportní procesy, které ovlivňují chování sinkové buňky?? Regulace související s transportními ději ?? Vztah zdroj - sink ?
Metabolizmus sinku Charakteristika sinku Síla sinku = aktivita sinku x velikost sinku • Metabolické sinky: apex, mladé, vyvíjející se listy, kořeny, vytvářející se plody, zásobní orgány Aktivita sinku závislá na : využití asimilátů pro potřeby růstua vývoje • Skladovací sinky: semena, hlízy, cibulky, skladovací kořeny, stonky, řapíky … Aktivita sinku závislá na :syntéze skladovacích látek – škrob, lipidy, proteiny U vyšších rostlin jsou sacharózasyntáza (SuSy) a invertázy klíčové enzymy regulace rozdělování sacharidů určují sílu sinku Metabolické sinky: převážně invertázy Skladovací sinky: převážně SuSy
Metabolizmus sinku Regulace aktivity sacharózasyntázy (SuSy) Exprese v sinku Exprese ve zdroji Exprese relativně nízká Exprese relativně vysoká Protein akumulovaný v etiolovaných listech degradován na světle během deetiolizace X fotosyntetické enzymy a SPS Degradace sacharózy Syntéza sacharózy Prudký pokles při tranzici sink- zdroj v listu V CC/SE komplexu – produkce energie pro dotování transportu ?? Produkce ADPG ..Syntéza škrobu ?? Několik genů s různou expresí (Arabidopsis 6 genů) – rozdíly v inhibici fruktózou Volná a asociovaná s membránou (účast na syntéze buněčné stěny) Př.:Transgenní brambor se sníženou expresí SuSy – snížení obsahu škrobu a snížení výnosu hlíz Př.:overexpreseSuSy u tabáku pod 35S promotorem --- zvýšení celkové biomasy
Metabolizmus sinku Regulace aktivity invertáz Velmi složitý systém regulace …. Důležitá funkce Regulace na transkripční a post-translační úrovni Problém .. Množství individuálních invertázových genů, které jsou různě regulovány. Všechny dosud studované rostliny mají 2 vakuolární invertázy ( VI) a několik stěnových invertáz (WI) (6 u Arabidopsis, 9 rýže). Regulace exprese závislá na pletivu • Regulace hormony (ABA, GA aktivitu) • Regulace cukry : většina indukovatelná sacharidy některé cukry reprimovatelné Posttranslační regulace • Regulace kinázami/fosfatázami (jak WI, VI), ale různé kinázy s různým regulačním potenciálem, interagující s ABA a cukernou signalizací a interakcí s patogeny. • Proteinové inhibitory – tvorba nefunkčního komplexu • Kompartmentace vakuolární invertázy do PPV (Precursor Protease Vesicles)
jádro plazmatická membrána WAK 2 Buněčná stěna PPV Metabolizmus sinku Regulace aktivity vakuolární invertázy invertáza VPE- proteáza aktivovatelná nízkým pH WAK: Wall-Associated Kinase • Kompartmentace vakuolární invertázy do PPV (Precursor Protease Vesicles, ER odvozené), následná fúze s tonoplastem, obsah vstupuje do vakuoly. Akce invertázy, postupná degradace nízkým pH aktivovanou proteázou – puls aktivity invertázy. • WAK2 ovlivňuje aktivitu vakuolární invertázy (nikoli stěnové), senzor turgoru, doména s kinázovou aktivitou v cytosolu. 22 členná WAK rodina u Arabidopsis. Huang et al., 2007
8 ječmen – výjimka ?? exprese ADPGázy špatný růst rostlin (nedostatek sacharózy pro rostoucí b.) exprese ADPGázy v hlízách zvýšení obsahu škrobu v hlízách exprese v hlízách snížení výnosu, prorůstání úžlab. pupenů, předčasné kvetení Metabolizmus sinku Syntéza škrobu ve zdroji a sinku Cytosolická ADPGaza ??? Sacharóza zdroj sink Je SuSy schopná produkovat ADPG, co to znamená ??? Trióza-P Pi ATP chloroplast amyloplast hexózy Pi ADP Trióza-P G-6-P Trióza-P Trióza-P/ Pitranslokátor G-6-P ATP/ADP translokátor G-1-P ADP-G/ADP transportér G-1-P Pi G-1-P Sacharóza SuSy?? ATP ATP G-6-P/Pi translokátor SuSy?? ADP-G ADP-G ADP-G ! ADP-G Některé rostliny dvě formy ADPGázy (SuSy??) Škrob ADP Škrob ADP Sacharóza Regulace syntézy : krátkodobá – allosterická inhibice/aktivace dlouhodobá – vývojové změny v expresi enzymů Transgenní rostliny : Martin Ludewig, 2007 Bahaji et al., 2011
Invertáza SuSy Metabolizmus sinku G-6-P G-1-P irreverzibilní reakce ?? PGM Poháněč syntézy škrobu ?? ? G-1P + ATP ADPG + PPi PPi2Pi Arabidopsis typically partitions 30–50% of its photoassimilates into starch ? PGM PPáza další poháněč syntézy škrobu ?? Lytovchenko et al., 2007 Fetke et al., 2011
Metabolizmus sinku Baroja-Fernandez et al., 2006 Příjem sacharózy endocytózou • Endocytické inhibitory : • sníží syntézu škrobu • sníží hladinu ADPG, • UDPG • neovlivní hladinu hexóza-P Žádný konflikt s transportem pomocí transportérů Žádný efekt inhibitorů při zásobování glukózou Acer pseudoplatanus (buněčné suspenze), disky hlíz bramboru, vyvíjející se endosperm ječmene Představa : počáteční fáze transportu (začátek dne) -- transportéry „vrcholná“ fáze transportu --- endocytóza
amyloplast vakuola cytosol apoplast Metabolizmus sacharidů ve skladovacím sinku Metabolizmus sinku Model čtyř hlavních „futile cycles „ Vykládání v plodech rajčete: 1.fáze :rychlý růst, sacharóza vykládána převážně symplastem,SuSy, degradace a resyntéza sacharózy (SuSy, SPS). (Veškerá import. sacharóza rozštěpena během 2 h). Vysoká akumulace škrobu, čistá akumulace škrobu 2.fáze :zpomalení růstu, sacharóza vykládána přes apoplast, degradace apoplastickou invertázou, resyntéza sacharózy v cytosolu, syntéza a degradace škrobu, čistá degradace škrobu Rychlost degradace škrobu konstantní, změna rychlosti syntézy změna čisté akumulace Kontrolní body: Aktivita ADPGázy Rychlost transportu G-6-P mezi amyloplastem a cytosolem • hydrolýzaSv apoplastu, transport hexóz do cytosolu a resyntéza sacharózy • kontinuální degradace SpůsobenímSuSya resyntézapomocí SuSyneboSPS v cytosolu ???Účel takto složitých vztahů ?? Efektivní kontrola hladin důležitých meziproduktů G-1-P; G-6-P; F-6-P; F,G, S Tím : regulace syntézy buněčné stěny, respirace, syntézy škrobu, skladování sacharózy, regulace osmotických poměrů • hydrolýza S ve vakuoleinvertázou, transport hexóz do cytosolu a resyntéza sacharózy • rychlá syntéza a degradace škrobu Futile = marný, zbytečný, neplodný, jalový Nguyen a Foyer, 2001
Metabolizmus sinku Když se sink stane zdrojem … • Srovnání degradace škrobu v listech, v dělohách klíčních rostlin a endospermu klíčících obilnin. • Listy : Degradace v chloroplastu, maltóza jako hlavní transportovaný sacharid do cytosolu. • Dělohy klíčních rostlin: Amyloplastová membrána je rozrušena, škrob je degradován cytosolickými enzymy. Glukanfosforyláza je důležitá při degradaci rozpustných glukanů. Sacharóza je syntetizována z degradovaného škrobu a transportována do kořenů a nadzemních částí semenáčků • Endosperm obilnin: Jak plastidová membrána, tak plazmalema jsou rozrušeny, škrob je degradován v neživých pletivech. Škrobová zrna jsou atakována -amylázou přeměněna na maltózu a glukózu. Glukóza a maltóza jsou exportovány z endospermu do štítku, a pak konvertovány na sacharózu, která podporuje růst embrya. Smith et al., 2005
Zdroj X sink Koncepce zdroje a sinku Zdroj: místo syntézy asimilátů Sink: místo utilizace Obecně: tok asimilátů od zdroje k sinku Zdroj: udává intenzitu transportu Sink: udává směr transportu Definovat pomocí: nabídka X poptávka Kombinace :fotoautotrofního, mixotrofního a heterotrofního způsobu výživy Jednoduchý vztah – klíční rostlina zdroj: dělohy sink: zejména kořen Situace se stává složitější s růstem a vývojem rostliny V čase dochází ke změnám Zdroj Sink…….. zakládání nových sinků, vyhasínání zdrojů….. Změna zdroj sink v rámci jednoho listu Neustálé znovuustavování rovnováhy zejména mezi syntézou asimilátů, spotřebou ve zdrojové buňce, transportem a utilizací v sincích X
Př.:Změny sink - zdroj během vývoje listu cukrovka – změny listu sink-zdroj asi při 40-50 % listové expanze exportující list SPS importující list 0 SPS Zdroj X sink Př.:Změna zdroj – sink během vývoje bramboru sink sink sink zdroj zdroj sink sink zdroj O morfologické změny, ale změny v genové expresi, proteinovém spektru, enzymatických aktivitách markery tranzice zdroj X sink Př.: Panašované listy – oblasti „zdrojové“ X oblasti „sinkové“ v jedné listové čepeli
Ovlivnění fytohormony Přímé ovlivnění : např. nakládání do floému: stimulace exogenním auxinem, inhibice exogenní ABA; opačný vliv exogenní aplikace na příjem v sinku Nepřímé ovlivnění : ovlivnění zakládání a růstu nových sinků. ovlivnění senescence, vývoje. Ovlivnění vztahů zdroj – sink Řízení síly sinku – komplexní záležitost vývojové signály (kvetení, tvorba zásobních orgánů.. ) signály o změnách prostředí (zastínění, nedostatek živin, abiotický i biotický stres) …… prostřednictvím hormonů (auxin, cytokininy, ABA, GA), signálních molekul, změn turgoru Př.:Ovlivnění síly sinku u cytokinin-deficientních rostlin Nadzemní část : obsahu chlorofylu, Ne - ovlivnění fotosyntetické kapacity a účinnosti, obsahu sacharidů síla zdroje nebyla ovlivněna. ALE sinková pletiva snížení obsahu NS sacharidů aktivity vakuolární invertázy obsahu ATP silné ovlivnění síly sinku Kořeny: mírné NS sacharidů, ALE : ovlivnění glykolýzy, obsahu ATP, a vývoje kořenů Regulace síly sinku cytokininem se liší v nadzemní části a kořenech
16 syntéza utilizace Př.:Celer: primární produkty fotosyntézy: sacharóza a manitol velmi mladý list 0 sacharóza rostoucí list sacharóza sacharóza, manitol sacharóza sacharóza manitol dospělý list Zdroj X sink Př.: odstranění plodů - změna alokace – často snížení rychlosti transportu – snížení rychlosti fotosyntézy Př.: zastínění listů, cukrovka, fazole Krátkodobé změny Zastavení růstu kořenů, přednostní zásobení mladých listů Dlouhodobé změny rychlosti fotosyntézy, aktivity RUBISCO, snížení obsahu škrobu, zvýšení obsahu sacharózy, transportu z listu, obnovení růstu kořenů.
manitol + NAD manóza + NADH +H+ manitoldehydrogenáza (MTD) manóza + ATP manóza-6-P + ADP hexokináza (HXK) manóza-6-P fruktóza-6-P fosfomanózaizomeráza Metabolizmus manitolu – ovlivnění stresem Metabolizmus sinku - Salinita , osmotický stres, nízká konc. Pi, vysoký obsah cukrů MTD + Napadení patogenem, vysoký obsah Pi, nízký obsah cukrů osmotický stres
zdroje Př.:Jabloň (ale i jiné dřeviny) jaro léto sink 1 sink 2 sink 3 ……… podzim růst kořenů růst listů růst kořenů Rozdělování mezi sinky výnosové parametry opylení růst výhonů Zakládání pupenů oplození apoplast: metabolizmus v apoplastu Závisí na účinnosti : • vykládání z floému • kompartmentaci • metabolické aktivitě Bujný růst plodů vede k omezení vývoje plodů následující rok kompetice Růst plodů symplast sink X zdroj spolupráce zdrojůXkompetice sinků Fitnes rostliny O alokaci rozhoduje relativní síla sinku Př.: Rozdělování asimilátů mezi sinky – ovlivnění výnosu - šlechtění bramboru : divoký typ: hlízy 7 % z celkové sušiny rostliny kulturní : 80% -„- X
Mechanismy podobné kompetici mezi geneticky totožnými částmi rostliny Př.: Solidago canadensis(Zlatobýl kanadský) Zastíněná část exportuje živiny (např. N) do části, která je může lépe využít
Růst prýtu… kompetice s ostatními rostlinami o světlo Krátkostébalné obilniny, + herbicidy potlačující konkurenční plevele Investice do reprodukčních struktur ….. množitelský potenciál Př.: Betula pubescens !!!Propojení dvou rostlin houbovými hyfami při mykorhize………..živiny ze zastíněné rostliny osvětlená rostlina stejného (nebo jiného) druhu nezískává Prýt X reprodukční struktury Komporomis Mechanismy, které v celku zvyšují životaschopnost (fitness) rostliny
Schema vlivu rhizobialní a AM symbiózy na fotosyntézu Voda, teplota, světlo, živiny CO2, Fotosyntéza Pool asimilátů Stimulace živinami Stimulace sinkem Náklady na růst semen Náklady na růst rhizobií Koncentrace N Náklady na růst prýtu Náklady na růst AM Koncentrace P Náklady na růst kořenů Rychlost fotosyntézy luskovin inokulovaných rhizobiálními a/nebo AM symbionty vzrostla oproti neinokulovaným rostlinám o 28% (rhizobiální), 14% (AM) a 51 % (kombinovananá inokulace)
Koordinace růstu Koordinace zásobování sacharidem a růstu rostliny X dostupnost zdroje C a energie během celého cyklu den-noc (mobilizace škrobu nebo jiných zásobních látek) Asimilace C jen na světle Jak toho dosáhnout Rovnováha mezi: asimilací C skladováním růstem ??? Pravděpodobná centrální role cukerné signalizace • Utilizace C se neřídí jednoduše momentálním přísunem asimilovaného C - „předvídání spotřeby během noci“ nutnost existence mechanismů řídících rychlost syntézy škrobu • Degradace škrobu během noci se děje lineárně, rychlostí „předvídající délku noci“ nutnost „měřit“ množství škrobu na konci dne • Změny podmínek prostředí vedou k „vyladění“ asimilace, skladování a spotřeby nutnost vnímání změn a přizpůsobení metabolizmu
Koordinace růstu noc den den den noc 12 h 12 h 6 h 18 h Syntéza a mobilizace škrobu Arabidopsis thaliana schopnost regulace až do 4 h fotoperiody Rostliny při režimu 12/12 malé změny v obsahu cukrů i růstu během denního cyklu Změna délky dnezměna rozdělování mezi škrob a rozpustné sacharidy ve dne změna rychlosti mobilizace během noci Př.: sója : 14 h fotoperioda … 60 % asimilátů škrob 7h -“- 90 % -“- • Rychle rostoucí genotypy Arabidopsis thaliana„více riskují“ – méně skladují a více získaného C investují do růstu Smith & Stitt, 2007; Stitt and Zeman, 2012
24 Koordinace růstu 3 4h Akutní odpověď: Náhlá změna; zastavení růstu Trvání změny ; vyladění rovnováhy mezi investicí do škrobu a mobilizací škrobu; obnovení růstu (s nižší rychlostí) Aklimační odpověď: Co se stane, když experimentálně náhle prodloužíme délku noci ? Syntéza škrobu a mobilizace škrobu • Dojde ke kompletnímu zastavení růstu, zejména kořenů noc den • Zastavení růstu trvá několik hodin po opětném vystavení světlu • Pokud tento režim trvá déle, dojde k ustavení nového rytmu syntézy škrobu a jeho utilizace v noci. 12h 12h Podobné přizpůsobení - reakce na sezónních změny, změny ozářenosti, teploty, dostupnosti vody…
Důsledky rozdílného podílu investice do růstu rostliny RGR - relativní růstová rychlost – zvýšení biomasy/ jednotku existující biomasy za den Dlouhodobější perspektiva mírně větší investice do růstu výrazně vyšší celková biomasa rostliny Stittand Zeeman, 2012
Koordinace růstu 4 12 h/12h 6 h/18h Genová exprese při nedostatku C Většina těchto genů vykazuje denní rytmus transkripce --- jakási příprava na nebezpečí nedostatku cukrů. Famine genes !!! Tato změna v trankriptech nevede ke změně hladiny enzymů Prodloužení noci vede ke změně transkripce během 4-6 hod, změny v aktivitě enzymů vyžadují několik dní. Vztahy mezi hladinami transkriptů a proteinů. Feast genes Jaké signály řídí denní změny v transkriptomu, a jak jsou spojeny s mechanismy koordinujícími C-stav a růst?? Kandidáti : cirkadienní rytmus, světlo a cukry 30% genů vykazuje denní rytmus transkripce v růžici Arabidopsis.
Koordinace růstu 4 12 h/12h 6 h/18h Genová exprese při nedostatku C Většina těchto genů vykazuje denní rytmus transkripce --- jakási příprava na nebezpečí nedostatku cukrů. Famine genes !!! Tato změna v trankriptech nevede ke změně hladiny enzymů Prodloužení noci vede ke změně transkripce během 4-6 hod, změny v aktivitě enzymů vyžadují několik dní. Vztahy mezi hladinami transkriptů a proteinů. Feast genes Jaké signály řídí denní změny v transkriptomu, a jak jsou spojeny s mechanismy koordinujícími C-stav a růst?? Kandidáti : cirkadienní rytmus, světlo a cukry 30% genů vykazuje denní rytmus transkripce v růžici Arabidopsis.
Koordinace růstu 4 12 h/12h 6 h/18h Famine genes Většina těchto genů vykazuje denní rytmus transkripce --- jakási příprava na nebezpečí nedostatku cukrů. !!! Tato změna v trankriptech nevede ke změně hladiny enzymů Prodloužení noci vede ke změně transkripce během 4-6 hod, změny v aktivitě enzymů vyžadují několik dní. Vztahy mezi hladinami transkriptů a proteinů. Feast genes Jaké signály řídí denní změny v transkriptomu, a jak jsou spojeny s mechanismy koordinujícími C-stav a růst?? Kandidáti : cirkadienní rytmus, světlo a cukry 30% genů vykazuje denní rytmus transkripce v růžici Arabidopsis.
SH HS Světlem indukovaná aktivace zprostředkovaná thioredoxinem Přizpůsobení syntézy škrobu po epizodě vyčerpání C Nedostatek G-6 -P SnRk1 dočasné zastavení růstu Ru-1,5 P2 Trióza-P Trióza-P Pi Zvýšení obsahu cukrů, když je obnovena dostupnost C syntéza T-6-P Fruktóza 1,6 bis-P Fruktóza 6-P Fruktóza 6-P SnRK1 Glukóza 6-P Glukóza 1-P Glukóza 1-P ATP ADPGáza PPi UDP-Glukóza fosfatáza ADP-Glukóza SPShigh SPSlow TRX SnRK1 Sacharóza 6-P Škrob Sacharóza T-6-P Cukrem indukovaná aktivace zprostředkovaná thioredoxinem (přes T-6-P) T-6-P cytosol chloroplast
Oxidace SH HS Aktivita transportérů Aktivita Aktivita ADPGazy Aktivita Oxidace Redukce Regulace redox stavem Lokalizace transportérů soustředěná do lipidických „raft-like„ domén. Zvýšení interakce protein-protein
Koordinace růstu 1 den tmy Regulace při nástupu reproduktivní fáze Náhlá epizoda sucha nebo teplotního stresu aborce semen v časném stadiu vývoje ! reakce na C hladovění, nikoli přímá reakce na stres samotný- předcházení nesplnitelným nárokům na přísun C při dalším vývoji semene semena založená „před stresem“ a „po stresu“ normálně pokračují ve vývoji Integrace dalších podnětů Síť interakcí je v reálu složitější: zahrnuje reakce na další podněty, jako je např. dostupnost živin nebo různá odpověď částí rostliny např. na signály o kvalitě světla Zahrnuje signalizaci mezi jednotlivými částmi rostliny, a tedy možnost vzájemného vyladění jejich rekce na změnu podmínek prostředí Předpoklad : cukry jsou významnou komponentou této signalizační sítě