Download
1 / 1
10 likes | 62 Views
Investigating DR6's role in apoptosis and NF-κB activation in different cell lines, shedding light on receptor functionality and shedding mechanisms.

E N D
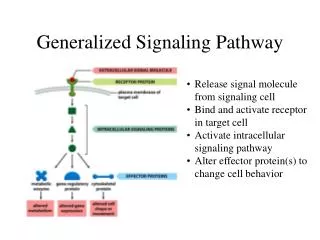
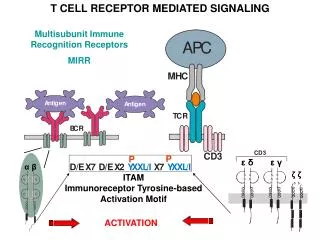
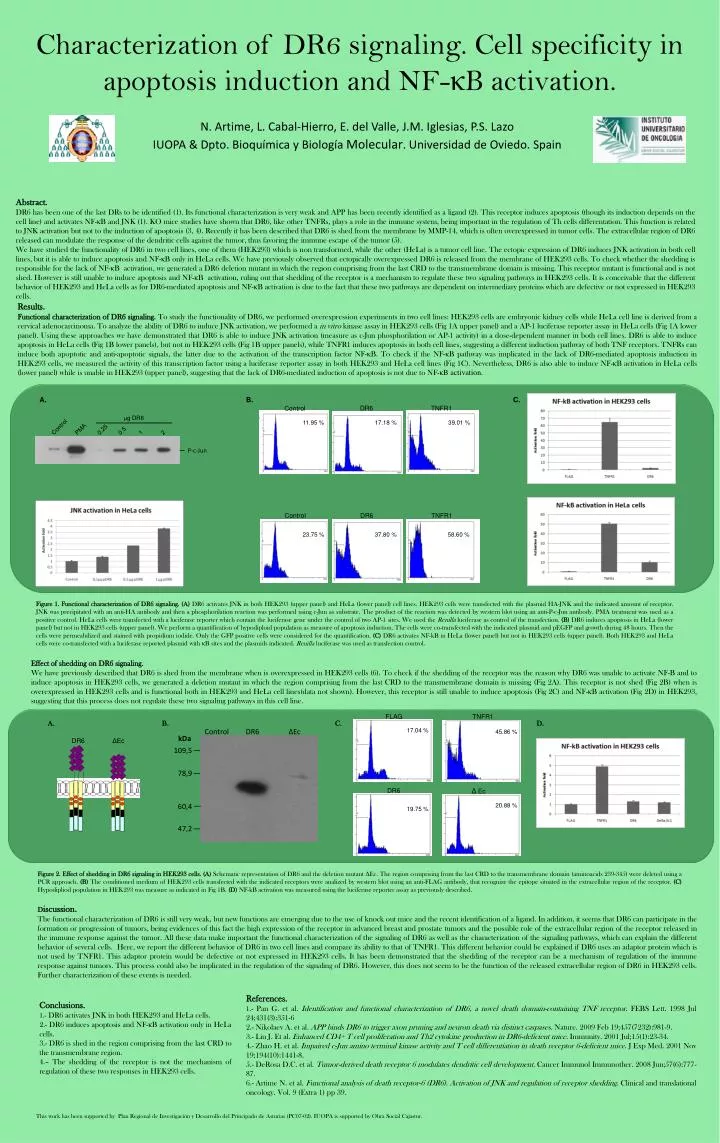
FLAG TNFR1 17.04 % 45.86 % DR6 Δ Ec 20.88 % 19.75 % Abstract. DR6 has been one of the last DRs to be identified (1). Its functional characterization is very weak and APP has been recently identified as a ligand (2). This receptor induces apoptosis (though its induction depends on the cell line) and activates NF-κB and JNK (1). KO mice studies have shown that DR6, like other TNFRs, plays a role in the immune system, being important in the regulation of Th cells differentation. This function is related to JNK activation but not to the induction of apoptosis (3, 4). Recently it has been described that DR6 is shed from the membrane by MMP-14, which is often overexpressed in tumor cells. The extracellular region of DR6 released can modulate the response of the dendritic cells against the tumor, thus favoring the immune escape of the tumor (5). We have studied the functionality of DR6 in two cell lines, one of them (HEK293) which is non transformed, while the other (HeLa) is a tumor cell line. The ectopic expression of DR6 induces JNK activation in both cell lines, but it is able to induce apoptosis and NF-κB only in HeLa cells. We have previously observed that ectopically overexpressed DR6 is released from the membrane of HEK293 cells. To check whether the shedding is responsible for the lack of NF-κB activation, we generated a DR6 deletion mutant in which the region comprising from the last CRD to the transmembrane domain is missing. This receptor mutant is functional and is not shed. However is still unable to induce apoptosis and NF-κB activation, ruling out that shedding of the receptor is a mechanism to regulate these two signaling pathways in HEK293 cells. It is conceivable that the different behavior of HEK293 and HeLa cells as for DR6-mediated apoptosis and NF-κB activation is due to the fact that these two pathways are dependent on intermediary proteins which are defective or not expressed in HEK293 cells. DR6 Control TNFR1 μg DR6 11.95 % 17.18 % 39.01 % Control PMA 0.25 0.5 1 2 P-c-Jun Results. Functional characterization of DR6 signaling. To study the functionality of DR6, we performed overexpression experiments in two cell lines: HEK293 cells are embryonic kidney cells while HeLa cell line is derived from a cervical adenocarcinoma. To analyze the ability of DR6 to induce JNK activation, we performed a in vitro kinase assay in HEK293 cells (Fig 1A upper panel) and a AP-1 luciferase reporter assay in HeLa cells (Fig 1A lower panel). Using these approaches we have demonstrated that DR6 is able to induce JNK activation (measure as c-Jun phosphorilation or AP-1 activity) in a dose-dependent manner in both cell lines. DR6 is able to induce apoptosis in HeLa cells (Fig 1B lower panels), but not in HEK293 cells (Fig 1B upper panels), while TNFR1 induces apoptosis in both cell lines, suggesting a different induction pathway of both TNF receptors. TNFRs can induce both apoptotic and anti-apoptotic signals, the latter due to the activation of the transcription factor NF-κB. To check if the NF-κB pathway was implicated in the lack of DR6-mediated apoptosis induction in HEK293 cells, we measured the activity of this transcription factor using a luciferase reporter assay in both HEK293 and HeLa cell lines (Fig 1C). Nevertheless, DR6 is also able to induce NF-κB activation in HeLa cells (lower panel) while is unable in HEK293 (upper panel), suggesting that the lack of DR6-mediated induction of apoptosis is not due to NF-κB activation. DR6 Control TNFR1 Characterization of DR6 signaling. Cell specificity in apoptosis induction and NF-κB activation. 23.75 % 37.80 % 58.60 % A. B. C. DR6 ΔEc Figure 1. Functional characterization of DR6 signaling.(A) DR6 activates JNK in both HEK293 (upper panel) and HeLa (lower panel) cell lines. HEK293 cells were transfected with the plasmid HA-JNK and the indicated amount of receptor. JNK was precipitated with an anti-HA antibody and then a phosphorilation reaction was performed using c-Jun as substrate. The product of the reaction was detected by western blot using an anti-P-c-Jun antibody. PMA treatment was used as a positive control. HeLa cells were transfected with a luciferase reporter which contain the luciferase gene under the control of two AP-1 sites. We used the Renilla luciferase as control of the transfection. (B) DR6 induces apoptosis in HeLa (lower panel) but not in HEK293 cells (upper panel). We perform a quantification of hypodiploid population as measure of apoptosis induction. The cells were co-transfected with the indicated plasmid and pEGFP and growth during 48 hours. Then the cells were permeabilized and stained with propidium iodide. Only the GFP positive cells were considered for the quantification. (C) DR6 activates NF-kB in HeLa (lower panel) but not in HEK293 cells (upper panel). Both HEK293 and HeLa cells were co-transfected with a luciferase reported plasmid with κB sites and the plasmids indicated. Renilla luciferase was used as transfection control. N. Artime, L. Cabal-Hierro, E. del Valle, J.M. Iglesias, P.S. Lazo IUOPA & Dpto. Bioquímica y Biología Molecular. Universidad de Oviedo. Spain Effect of shedding on DR6 signaling. We have previously described that DR6 is shed from the membrane when is overexpressed in HEK293 cells (6). To check if the shedding of the receptor was the reason why DR6 was unable to activate NF-B and to induce apoptosis in HEK293 cells, we generated a deletion mutant in which the region comprising from the last CRD to the transmembrane domain is missing (Fig 2A). This receptor is not shed (Fig 2B) when is overexpressed in HEK293 cells and is functional both in HEK293 and HeLa cell lines(data not shown). However, this receptor is still unable to induce apoptosis (Fig 2C) and NF-κB activation (Fig 2D) in HEK293, suggesting that this process does not regulate these two signaling pathways in this cell line. A. B. C. D. Control DR6 ΔEc kDa 109,5 78,9 60,4 47,2 Figure 2. Effect of shedding in DR6 signaling in HEK293 cells. (A) Schematic representation of DR6 and the deletion mutant ΔEc. The region comprising from the last CRD to the transmembrane domain (aminoacids 239-345) were deleted using a PCR approach. (B) The conditioned medium of HEK293 cells transfected with the indicated receptors were analized by western blot using an anti-FLAG antibody, that recognize the epitope situated in the extracellular region of the receptor. (C) Hypodipliod population in HEK293 was measure as indicated in Fig 1B. (D) NF-kB activation was measured using the luciferase reporter assay as previously described. Discussion. The functional characterization of DR6 is still very weak, but new functions are emerging due to the use of knock out mice and the recent identification of a ligand. In addition, it seems that DR6 can participate in the formation or progression of tumors, being evidences of this fact the high expression of the receptor in advanced breast and prostate tumors and the possible role of the extracellular region of the receptor released in the immune response against the tumor. All these data make important the functional characterization of the signaling of DR6 as well as the characterization of the signaling pathways, which can explain the different behavior of several cells. Here, we report the different behavior of DR6 in two cell lines and compare its ability to that of TNFR1. This different behavior could be explained if DR6 uses an adaptor protein which is not used by TNFR1. This adaptor protein would be defective or not expressed in HEK293 cells. It has been demonstrated that the shedding of the receptor can be a mechanism of regulation of the immune response against tumors. This process could also be implicated in the regulation of the signaling of DR6. However, this does not seem to be the function of the released extracellular region of DR6 in HEK293 cells. Further characterization of these events is needed. References. 1.- Pan G. et al. Identification and functional characterization of DR6, a novel death domain-containing TNF receptor. FEBS Lett. 1998 Jul 24;431(3):351-6 2.- Nikolaev A. et al. APP binds DR6 to trigger axon pruning and neuron death via distinct caspases. Nature. 2009 Feb 19;457(7232):981-9. 3.- Liu J. Et al. Enhanced CD4+ T cell proliferation and Th2 cytokine production in DR6-deficient mice. Immunity. 2001 Jul;15(1):23-34. 4.- Zhao H. et al. Impaired c-Jun amino terminal kinase activity and T cell differentiation in death receptor 6-deficient mice. J Exp Med. 2001 Nov 19;194(10):1441-8. 5.- DeRosa D.C. et al. Tumor-derived death receptor 6 modulates dendritic cell development. Cancer Immunol Immunother. 2008 Jun;57(6):777-87. 6.- Artime N. et al. Functional analysis of death receptor-6 (DR6). Activation of JNK and regulation of receptor shedding. Clinical and translational oncology. Vol. 9 (Extra 1) pp 39. Conclusions. 1.- DR6 activates JNK in both HEK293 and HeLa cells. 2.- DR6 induces apoptosis and NF-κB activation only in HeLa cells. 3.- DR6 is shed in the region comprising from the last CRD to the transmembrane region. 4.– The shedding of the receptor is not the mechanism of regulation of these two responses in HEK293 cells. This work has been supported by Plan Regional de Investigación y Desarrollo del Principado de Asturias (PC07-02). IUOPA is supported by Obra Social Cajastur.