Download
1 / 1
10 likes | 94 Views
Testing the r' method of estimating per capita growth rate in Aedes albopictus Matthew Chmielewski, Camilo Khatchikian and Todd Livdahl Department of Biology, Clark University, Worcester MA. Abstract

E N D
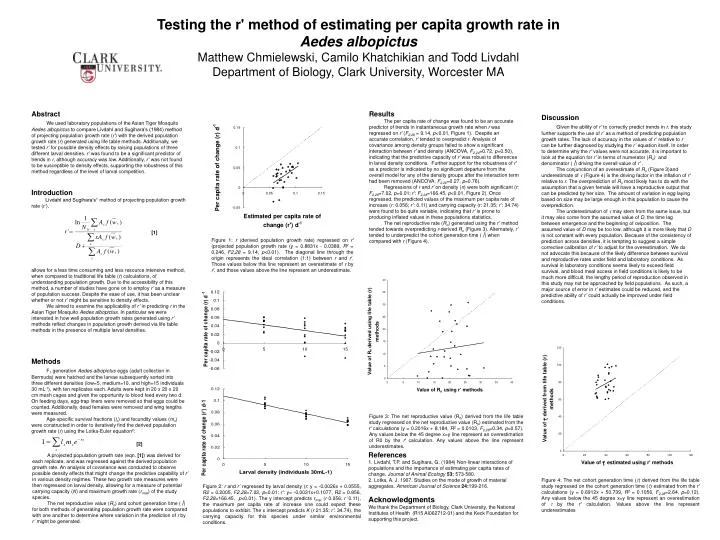
Testing the r' method of estimating per capita growth rate inAedes albopictusMatthew Chmielewski, Camilo Khatchikian and Todd LivdahlDepartment of Biology, Clark University, Worcester MA Abstract We used laboratory populations of the Asian Tiger Mosquito Aedes albopictus to compare Livdahl and Sugihara’s (1984) method of projecting population growth rate (r') with the derived population growth rate (r) generated using life table methods. Additionally, we tested r’ for possible density effects by raising populations of three different larval densities. r’ was found to be a significant predictor of trends in r, although accuracy was low. Additionally, r’ was not found to be susceptible to density effects, supporting the robustness of this method regardless of the level of larval competition. Results The per capita rate of change was found to be an accurate predictor of trends in instantaneous growth rate when r was regressed on r′ (F2,28 = 9.14, p<0.01, Figure 1). Despite an accurate correlation, r’ tended to overpredict r. Analysis of covariance among density groups failed to show a significant interaction between r′ and density (ANCOVA, F2,24=0.72, p=0.50), indicating that the predictive capacity of r′ was robust to differences in larval density conditions. Further support for the robustness of r′ as a predictor is indicated by no significant departure from the overall model for any of the density groups after the interaction term had been removed (ANCOVA, F2,26=0.27, p=0.76). Regressions of r and r′ on density (n) were both significant (r: F2,28=7.02, p=0.01; r′: F2,28=166.45, p<0.01, Figure 2). Once regressed, the predicted values of the maximum per capita rate of increase (r: 0.056; r’:0.11) and carrying capacity (r: 21.35; r’: 34.74) were found to be quite variable, indicating that r’ is prone to producing inflated values in these populations statistics. The net reproductive rate (Ro) generated using the r’ method tended towards overpredicting r-derived Ro(Figure 3). Alternately, r’ tended to underpredict the cohort generation time (τ) when compared with r (Figure 4). Discussion Given the ability of r’ to correctly predict trends in r, this study further supports the use of r’ as a method of predicting population growth rates. The lack of accuracy in the values of r’ relative to r can be further diagnosed by studying the r’ equation itself. In order to determine why the r′ values were not accurate, it is important to look at the equation for r′ in terms of numerator (Ro) and denominator (τ) driving the overall value of r′ . The conjunction of an overestimate of Ro (Figure 3)and underestimate of t (Figure 4) is the driving factor in the inflation of r′ relative to r. The overprediction of Ro most likely has to do with the assumption that a given female will have a reproductive output that can be predicted by her size. The amount of variation in egg laying based on size may be large enough in this population to cause the overprediction. The underestimation of t may stem from the same issue, but it may also come from the assumed value of D, the time lag between emergence and the beginning of oviposition. The assumed value of D may be too low, although it is more likely that D is not constant with every population. Because of the consistency of prediction across densities, it is tempting to suggest a simple corrective calibration of r’ to adjust for the overestimation. We do not advocate this because of the likely difference between survival and reproductive rates under field and laboratory conditions. As survival in laboratory conditions seems likely to exceed field survival, and blood meal access in field conditions is likely to be much more difficult, the lengthy period of reproduction observed in this study may not be approached by field populations. As such, a major source of error in r’ estimates could be reduced, and the predictive ability of r’ could actually be improved under field conditions. Introduction Livdahl and Sugihara’s1 method of projecting population growth rate (r’), [1] Figure 1: r (derived population growth rate) regressed on r′ (projected population growth rate (y = 0.8831x - 0.0389, R2 = 0.246, F2,28 = 9.14, p<0.01). The diagonal line through the origin represents the ideal correlation (1:1) between r and r′. Those values below this line represent an overestimate of r by r′, and those values above the line represent an underestimate. allows for a less time consuming and less resource intensive method, when compared to traditional life table (r) calculations, of understanding population growth. Due to the accessibility of this method, a number of studies have gone on to employ r’ as a measure of population success. Despite the ease of use, it has been unclear whether or not r’ might be sensitive to density effects. We aimed to examine the applicability of r’ in predicting r in the Asian Tiger Mosquito Aedes albopictus. In particular we were interested in how well population growth rates generated using r’ methodsreflect changes in population growth derived via life table methods in the presence of multiple larval densities. Methods F1 generation Aedes albopictus eggs (adult collection in Bermuda) were hatched and the larvae subsequently sorted into three different densities (low=5, medium=10, and high=15 individuals 30 mL-1), with ten replicates each. Adults were kept in 20 x 20 x 20 cm mesh cages and given the opportunity to blood feed every two d. On feeding days, egg-trap liners were removed so that eggs could be counted. Additionally, dead females were removed and wing lengths were measured. Age-specific survival fractions (lx) and fecundity values (mx) were constructed in order to iteratively find the derived population growth rate (r) using the Lotka-Euler equation2: Figure 3: The net reproductive value (R0) derived from the life table study regressed on the net reproductive value (R0) estimated from the r′ calculations (y = 0.2016x + 8.184, R2 = 0.0103, F2,28=0.34, p=0.57). Any values below the 45 degree x=y line represent an overestimation of R0 by the r′ calculation. Any values above the line represent underestimates. [2] References 1. Livdahl, T.P. and Sugihara, G. (1984) Non-linear interactions of populations and the importance of estimating per capita rates of change. Journal of Animal Ecology 53: 573-580. 2. Lotka, A. J. 1907. Studies on the mode of growth of material aggregates. American Journal of Science24:199-216. Acknowledgments We thank the Department of Biology, Clark University, the National Institutes of Health (R15 AI062712-01)and the Keck Foundation for supporting this project. A projected population growth rate (eqn. [1]) was derived for each replicate, and was regressed against the derived population growth rate. An analysis of covariance was conducted to observe possible density effects that might change the predictive capability of r’ in various density regimes. These two growth rate measures were then regressed on larval density, allowing for a measure of potential carrying capacity (K) and maximum growth rate (rmax) of the study species. The net reproductive value (Ro) and cohort generation time (τ) for both methods of generating population growth rate were compared with one another to determine where variation in the prediction of r by r’ might be generated. Figure 4: The net cohort generation time (t) derived from the life table study regressed on the cohort generation time (t) estimated from the r′ calculations (y = 0.6912x + 50.739, R2 = 0.1056, F2,28=2.64, p=0.12). Any values below the 45 degree x=y line represent an overestimation of t by the r′ calculation. Values above the line represent underestimates Figure 2: r and r’ regressed by larval density (r: y = -0.0026x + 0.0555, R2 = 0.2005, F2,28=7.02, p=0.01; r’: y= -0.0031x+0.1077, R2 = 0.856, F2,28=166.45, p<0.01). The y intercept predicts rmax (r 0.056; r’ 0.11), the maximum per capita rate of increase one could expect these populations to exhibit. The x intercept predicts K (r 21.35; r’: 34.74), the carrying capacity for this species under similar environmental conditions.