Download
1 / 1
10 likes | 132 Views
Cardiac Bioelectricity and Arrhythmia Center (CBAC). A computational model of β-adrenergic signaling in cardiac myocytes: Local signaling and rate-dependent action-potential effects. Jordi Heijman (1,2) , Paul G.A. Volders (2) , Ronald L. Westra (2) and Yoram Rudy (1).

E N D
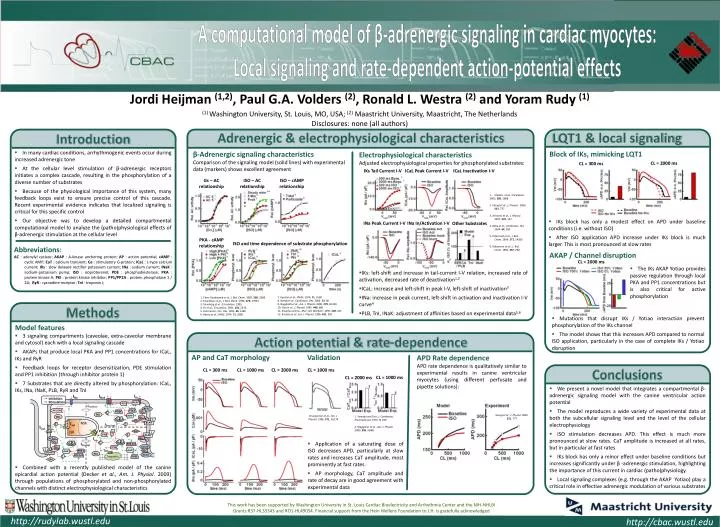
Cardiac Bioelectricity and Arrhythmia Center (CBAC) A computational model of β-adrenergic signaling in cardiac myocytes: Local signaling and rate-dependent action-potential effects Jordi Heijman (1,2), Paul G.A. Volders (2), Ronald L. Westra (2) and Yoram Rudy (1) (1) Washington University, St. Louis, MO, USA; (2) Maastricht University, Maastricht, The Netherlands Disclosures: none (all authors) Adrenergic & electrophysiological characteristics LQT1 & local signaling Block of IKs, mimicking LQT1 AKAP / Channel disruption Introduction • In many cardiac conditions, arrhythmogenic events occur during increased adrenergic tone • At the cellular level stimulation of β-adrenergic receptors initiates a complex cascade, resulting in the phosphorylation of a diverse number of substrates • Because of the physiological importance of this system, many feedback loops exist to ensure precise control of this cascade. Recent experimental evidence indicates that localized signaling is critical for this specific control • Our objective was to develop a detailed compartmental computational model to analyze the (patho)physiological effects of β-adrenergic stimulation at the cellular level β-Adrenergic signaling characteristics Comparison of the signaling model (solid lines) with experimental data (markers) shows excellent agreement • Electrophysiological characteristics • Adjusted electrophysiological properties for phosphorylated substrates: • IKs: left-shift and increase in tail-current I-V relation, increased rate of activation, decreased rate of deactivation1,2 • ICaL: increase and left-shift in peak I-V, left-shift of inactivation3 • INa: increase in peak current, left-shift in activation and inactivation I-V curve4 • PLB, TnI, INaK: adjustment of affinities based on experimental data5,6 CL = 2000 ms CL = 300 ms IKs Tail Current I-V ICaL Peak Current I-V ICaL Inactivation I-V Gs – AC relationship ISO – AC relationship ISO – cAMP relationship • Volders et al., Circulation • 2003. 280, 1808 • 2. Stengl et al., J. Physiol. 2003. 551, 777 • 3. Antoons et al., J. Physiol. 2007. 579, 147 • 4. Baba et al., Cardiovasc. Res. 2004. 64, 260 • 5. Odermatt et al., J. Biol. Chem. 1996. 271, 14206 • 6. Robertson et al., J. Biol. Chem. 1982. 257, 260 • IKs block has only a modest effect on APD under baseline conditions (i.e. without ISO) • After ISO application APD increase under IKs block is much larger. This is most pronounced at slow rates INa Peak Current I-V INa In/Activation I-V Other Substrates PKA - cAMP relationship ISO and time dependence of substrate phosphorylation Abbreviations: AC : adenylyl cyclase; AKAP : A-kinase anchoring protein; AP : action potential; cAMP : cyclic AMP; CaT : calcium transient; Gs : stimulatory G-protein; ICaL : L-type calcium current; IKs : slow delayed rectifier potassium current; INa : sodium current; INaK : sodium-potassium pump; ISO : isoproterenol; PDE : phosphodiesterase; PKA : protein kinase A; PKI : protein kinase inhibitor; PP1/PP2A : protein phosphatase 1 / 2A; RyR : ryanodine receptor; TnI : troponin I; CL = 2000 ms • The IKs AKAP Yotiao provides passive regulation through local PKA and PP1 concentrations but is also critical for active phosphorylation 7. Kuschel et al., PNAS. 1974. 71, 3580 8. Stengl et al., Cardiovasc. Res. 2006. 72, 90 9. Nagykaldi et al., Am. J. Physiol. 2000. 279, H1329 10. Gao et al., J. Physiol. 1992. 449, 689 11. Sulakhe and Vo., Mol. Cell. Biochem. 1995. 149, 103 12. Kirstein et al., Eur. J. Physiol. 1996. 431, 395 1. Chen-Goodspeed et al., J Biol. Chem. 2005. 280, 1808 2. Freedman et al., J. Biol. Chem. 1996. 270, 17953 3. Steinberg et al., Circulation. 1995. 4. Yu et al., Circulation. 2001. 102, 2535 5. Hohl and Li, Circ. Res. 1991. 69, 1369 6. Beavo et al., PNAS. 1974. 71, 3580 • Mutations that disrupt IKs / Yotiao interaction prevent phosphorylation of the IKs channel • The model shows that this increases APD compared to normal ISO application, particularly in the case of complete IKs / Yotiao disruption Action potential & rate-dependence AP and CaT morphology Validation APD Rate dependence APD rate dependence is qualitatively similar to experimental results in canine ventricular myocytes (using different perfusate and pipette solutions): • Model features • 3 signaling compartments (caveolae, extra-caveolar membrane and cytosol) each with a local signaling cascade • AKAPs that produce local PKA and PP1 concentrations for ICaL, IKs and RyR • Feedback loops for receptor desensitization, PDE stimulation and PP1 inhibition (through inhibitor protein 1) • 7 Substrates that are directly altered by phosphorylation: ICaL, IKs, INa, INaK, PLB, RyR and TnI • Combined with a recently published model of the canine epicardial action potential (Decker et al., Am. J. Physiol. 2009) through populations of phosphorylated and non-phosphorylated channels with distinct electrophysiological characteristics • We present a novel model that integrates a compartmental β-adrenergic signaling model with the canine ventricular action potential • The model reproduces a wide variety of experimental data at both the subcellular signaling level and the level of the cellular electrophysiology • ISO stimulation decreases APD. This effect is much more pronounced at slow rates. CaT amplitude is increased at all rates, but in particular at fast rates • IKs block has only a minor effect under baseline conditions but increases significantly under β -adrenergic stimulation, highlighting the importance of this current in cardiac (patho)physiology. • Local signaling complexes (e.g. through the AKAP Yotiao) play a critical role in effective adrenergic modulation of various substrates Conclusions Methods CL = 300 ms CL = 1000 ms CL = 2000 ms CL = 1000 ms CL = 1000 ms CL = 2000 ms Stengl et al., J. Physiol. 2003. 551, 777 Charpentier et al., Am. J. Physiol. 1996. 271, H1174 1. Yamada and Corr, J. Cardiovasc. Electrophysiol. 1992. 3, 209 2. Waggoner et al. ,Am. J. Physiol. 2009. 296, H698 • Application of a saturating dose of ISO decreases APD, particularly at slow rates and increases CaT amplitude, most prominently at fast rates. • AP morphology, CaT amplitude and rate of decay are in good agreement with experimental data This work has been supported by Washington University in St. Louis Cardiac Bioelectricity and Arrhythmia Center and the NIH-NHLBI Grants R37-HL33343 and RO1-HL49054. Financial support from the Hein Wellens Foundation to J.H. is gratefully acknowledged http://rudylab.wustl.edu http://cbac.wustl.edu