Download
1 / 50
500 likes | 691 Views
kokkusobivus teoreetilise jaotusega. A. Rogers & H. Harpendig. 30k. populatsiooni suuruse fluktueerumine. Kui populatsiooni suurus kahaneb on võimalik Koalestsentsiaegu korrigeerida:. 60k -> 50k + 10x10=150k. N. N/10. 0. 50k. 100k. Avise 2000. tähekujulisus. =1 Keskmine mutat-

E N D
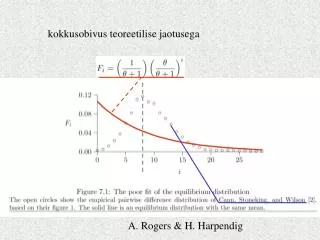
kokkusobivus teoreetilise jaotusega A. Rogers & H. Harpendig
30k populatsiooni suuruse fluktueerumine Kui populatsiooni suurus kahaneb on võimalik Koalestsentsiaegu korrigeerida: 60k -> 50k + 10x10=150k N N/10 0 50k 100k Avise 2000
tähekujulisus =1 Keskmine mutat- sioonide arv puul rajajatüübini pop size expansion time mismatch distribution =2 Koalestsentsiaja hindamine Forster et al. 1996 =/ =2=s=
Tajima D Neutraalsuse korral: D=0 Kergelt kahjulikke mutatsioone hoitakse madala sageduse juures; D<0 positiivne valik, vastupidiselt D>0
coalescence "THEORIE DE LA COALESCENCE" Kingman, Griffiths, Tavare, Hudson
coalescence D~0 D<0 D>0 D sõltub puu kujust, vastavalt selektsioonile, populatsiooni suuruse Muutustele jm.
Jorde et al. 1995 Populatsioonide vaheline paariviisiline mittekokkulangemine
5 x 1 = 5 1 x 2 = 2 2 x 3 = 6 8 13 = 1.625 mismatch distribution 10 x 2 = 20 5 x 3 = 15 10 x 4 = 40 2 x 5 = 10 1 x 6 = 6 91 =91/28 =3.25 /2= 1.625 s=10/2.715= 3.68 Tähekujulise puu korral isegi kui jaotus pole unimodaalne võrdub 2 ja s>
pop suurus ekspansioon aeg Tähekujuline puu 5 x 0 8 x 1 = 8 13 8 rho=8/13=0.615 5 mismatch distribution =1.23 =2 kõik 78 paariviisilist võrdlust 78-st Koalestseeruvad samal rajajatüübil: täheindeks = 1 Torroni et al. 1998
täheindeks 27 paariviisilist võrdlust 28-st Koalestseeruvad samal rajajatüübil: täheindeks = 0.96
Oodatud erinevuste arv Poissoni jaotuse järgi tähetopoloogia korral
28 paari 1770st ei koalestseeru CRS-il; index =0,984 11 x 0 22 x 1 = 22 16 x 2 = 32 9 x 3 = 27 1 x 4 = 4 --------------- 59 85 rho=1.44 näide 2=4,216 f=5 p=0,52 - mitte oluliselt erinev
29 x 0 88 x 1 = 88 55 x 2 = 110 16 x 3 = 48 3 x 4 = 12 ----------------- 191 258 rho=1.35 2650 paari 18145st ei koalestseeru Rajajal, puuon 0,85 star like 2=12,494 f=5 p=0,029 - oluline 95% juures
in star topology, even if distribution is not unimodal equals 2 and s>
star index Both: rho=1 X2=0 (with Poisson) differ by star index
and s do not will estimate coalescence time c b a 8 tips -> 8x7/2=28 pairs 4 x 2 = 8 8 x 4 = 32 16x 6 = 96 136 =136/28 =4.85 /2= 2.4 0 x 0 0 x 1 0 x 2 8 x 3 = 24 8 24 = 3 s=14/2,715=5,16 s/2=2,58
c b a 8 tips -> 8x7/2=28 pairs 4 x 2 = 8 8 x 4 = 32 16x12 = 192 232 =232/28 =8.29 /2= 2.4 0 x 0 0 x 1 0 x 2 8 x 6 = 48 8 48 = 6 s=20/2,715=7,36
summing up , S are indirect in (tree is not needed for) estimating the coalescent , S are not reliable in estimating coalescence times =2 only in star phylogeny can be used to estimate coalescence times given the tree the variance of a coalescent estimate depends on tree topology
when tree is not star like, the variance of rho is estimated as (Saillard et al. 2000): rho V(rho) t=rho V
2 1x 1 = 1 1 1 2 1x (1+1) = 4 2 1 2 3 = 9 1x 1x 3=9 2 2=8 2x 1 2 1=2 2x when tree is not star like, the variance of rho can be estimated as (Saillard et al. 2000): rho V(rho) 1 1 t= u uV 1 E=26/16=1.625 V=66/256, V= 0.51 2 4 1 3 1 1 1 25 1 2 1
fülogeograafia ja ökogeograafia biogeograafia tegeleb organismide levikuga ökogeograafia valdkond on ökoloogilises, fülogeograafial geoloogilises ajaskaalas ökogeograafia keskendub kaasajale ja uurib valiku, populatsiooni dünaamika ning keskkonna toimet populatsiooni struktuurile fülogeograafia, omades ka laiemat geograafilist haaret, otsib seoseid geoloogiliste protsesside (laamtektoonika, kliimamuutused, merevee taseme muutused) ja fülogeneesi (geenipuud) vahel. Levik, migratsioon (dispersal), tuumalad (centres of origin) ja varjupaigad (refugia) on põhilised seletused. nende kahe eristamise mõtteks on ajalise ja geograafilise perspektiivi rõhutamine
Peromyscus hiire MP puu kujutamine vastavalt mtDNA haplotüüpide geograafilisele levikule
alampopulatsioonide levik ja eristumine geograafiline kliin (alleelisageduste sujuv kauguseline üleminek) barjäär aeg Croizat (1964) dünaamiline koevolutsioon maa ja tema biosfääri vahel vikaarsus: eri liikidel samas geograafilises kontekstis samasugune fülogeograafiline jaotumus. Geograafiliselt diskreetsete alleelisageduste kujunemine eri liikidel viitab ühisele põhjusele. levimise (dispersal) korral tekivad erinevused alampopulatsioonide vahel algpopulatsioonist migratsiooni ja sellele järgneva triivi tulemusena.
mõisted gene flow(geenimigratsioon) - alleelide liikumine (indiviidide kaudu) liigi alampopulatsioonide vahel isolation-by-distance (isoleeritus kauguse läbi) - geneetilise kauguse suurenemine joones geograafilise kaugusega range expansion (levikuline ekspansioon) - ekspansiooniga kaasnev leviala laeinemine vicariance (vikaarsus) - populatsiooni ajaloos toimuv protsess, mille käigus levila jaotumine (n. barjääri kaudu) viib alampopulatsioonide geograafilise isoleerumiseni. Alleelisageduste erinevus tekkivates alampopulatsioonides on määratud ennekõike eelnevalt eksiteerunud populatsiooni alamstruktuuri poolt (isoleeritus kauguse läbi, kliinid jm.) Croizat: 'centres of origin' -> 'generalized tracks' Avise: kladistiline fülogeograafia: taksonite ja levilate fülogeograafia kokkulangemine
kliima 20,000 a. tagasi Hewitt Nature 405:907-913 (2000)
Euroopa hübriidtsoonid
Inimese Y kromosoomi geograafilised barjäärtsoonid Rosser et al. 2000 AmJHumGenet
populatsiooni demograafiline ajalugu ja fülogeograafia koalestsentsiteooria Markovi ahel (Markov chain) - ahel, milles üleminek ühelt seisundilt teisele ei sõltu 'seisundite ajaloost' ehk millises seisundis oli üleelmine ahela lüli branching-process,demograafiline mudel, mille käigus iga populatsiooni osake toodab järelkasvu teistest osakestest sõltumatult ja vastavalt Markovi ahelale põlvkonnast põlvkonda liinide sorteerumine (lineage sorting) - tõenäosuslik protsess, mille käigus vastavalt perekonna suuruste juhuslikule jaotusele (Poisson) mingi osa liinidest populatsioonis säilub ning teised surevad välja
liinide sorteerumine 7 liinist 1 7 liinist 1 ema- ja isaliinide ajalood on analoogsed aga mitte identsed
liinide kaotsimineku tõenäosused Näiteks, populatsioonis, kus emad sünnitavad tütreid vastavalt Poissoni jaotusele keskmisega (seega ka varieeruvusega) 1.0 (), on tütardeta emade tõenäosuseks e-1=0.37, ja 1, 2, 3 ja 4 tütrega emade sageduseks (tõenäosuseks) 0.37, 0.18, 0.06 ja 0.01, vastavalt. Seega, ühe põlvkonna jooksul läheb konstantse suurusega (=1) populatsioonis kaotsi 36.8% emaliinidest.
liinide kaotsiminek n põlvkonna jooksul p - kaotsiminek q - allesjäämine pN - populatsiooni kadumine emaliinide kaotsiminekuga - enam kui kahe liini ellujäämine cb - enam kui kahe liini ellujäämine tingimusel, et populatsioon ei surnud välja populatsioonis, kus on näiteks 10,000 emast, jääb ligilähedaseks ühele esimesed 10,000 põlvkonda ja kahaneb nulli lähedale 100,000 põlvkonna jooksul; kui N on 10, siis koalestseeruvad liinid peaaegu kindlasti 100 põlvkonna jooksul.
kumuleeruv liinide ellujäämise tõenäosus (q) sõltuvuses populatsiooni suurusest
kumulatiivsed tõenäosused kahe või enama liini säilu- miseks konstantse suurusega populatsioonis suurusega 2 kuni 10,000 liini
liinide säilumine kasvavas ja kahanevas populatsioonis
kaotsimineku tõenäosused sõltuvalt keskimisest perekonna suurusest põlvkonnad
populatsiooni kasv sõltuvalt keskmisest järeltulijate arvust näo kohta
mitokondriaalse DNA varieeruvus mustlastel
mitokondriaalse DNA varieeruvus mustlastel , cb,ligilähedased ühele
reproduktiivse edukuse varieeruvus suurem varieeruvus kiirendab liinide sorteerumist ja koalestsentsi aeg väheneb
koalestsents demograafilised faktorid: a) varieeruvus järglaste arvus b) fluktuatsioonid c) põlvkondade kattuvus arvesse võetavad Ne hindamisel olulisem kaal populatsiooni ajaloo varaseimas järgus toimunul effektiivne populatsiooni suurus annab genealoogilise sügavuse, kuid fluktuatsioonid jätavad jälje puu topoloogiasse, nn. ajaline klasterdumine (temporal clustering)
fluktuatsioonid populatsiooni demograafilises ajaloos ekspansioon <> pudelikael lained paariviisiliste erinevuste jaotustes (waves in mismatch distributions) MMD aga hindab erinevusi üle kogu puu Nee et al. 1996: tuleb keskenduda puule ja lokaalselt iga harunemist eraldi käsitleda, plottida liinide arv puul vastavalt harunemiste sügavusele -> A - konstantne populatsioon B - kasvav populatsioon, tõusunurk kasvukiirus ent, töötab vaid ultrameetrilise puu korral, vähem robustne kui MMD, ei arvesta harude varieeruvust, populatsiooni alamstruktuuri rohkem visuaalne kui täppismeetod
fluktuatsioonid populatsiooni demograafilises ajaloos madal h ja madal p: hiljutine tugev pudelikael, selektsioon vm. kõrge h ja kõrge p: stabiilne pikaajaline populatsioon kõrge Ne-ga, või siis tugev segunemine kõrge h ja madal p: kiire kasv madala Ne-ga populatsioonist suhteliselt hiljuti madal h ja kõrge p: pudelikaela seisund suurest eellaspopulatsioonist lähtuvalt; segunemine kahe väikse, geogr jaotunud alampop vahel kuigi neid statistikuid on kerge arvutada, ei ütle nad konkreetselt palju populatsiooni ajaloo kohta
populatsioonide ruumiline struktuur isoleeritud populatsioonid puhverdab liinide sorteerumist ja pikendab totaalset koalestsentsi liigis, mis on koosnenud G põlvkonna jooksul mitmest isoleeritud populatsioonist, on minimaalne koalestsents määratud G poolt bimodaalne MM ja IM -> isoleeritud alampopulatsioonides sisaldub ka lahknemisele eelnenud varieeruvus, selle korrigeerimiseks
min tAB max tAA ruumiline struktuur: genealoogiline lähenemine liinide sorteerumise tulemusena võimalikud kolme tüüpi topoloogiad: distantsarvutuslikult:
inimese polüfüleetiline mitokondripuu Euraasias

