Download
1 / 1
20 likes | 206 Views
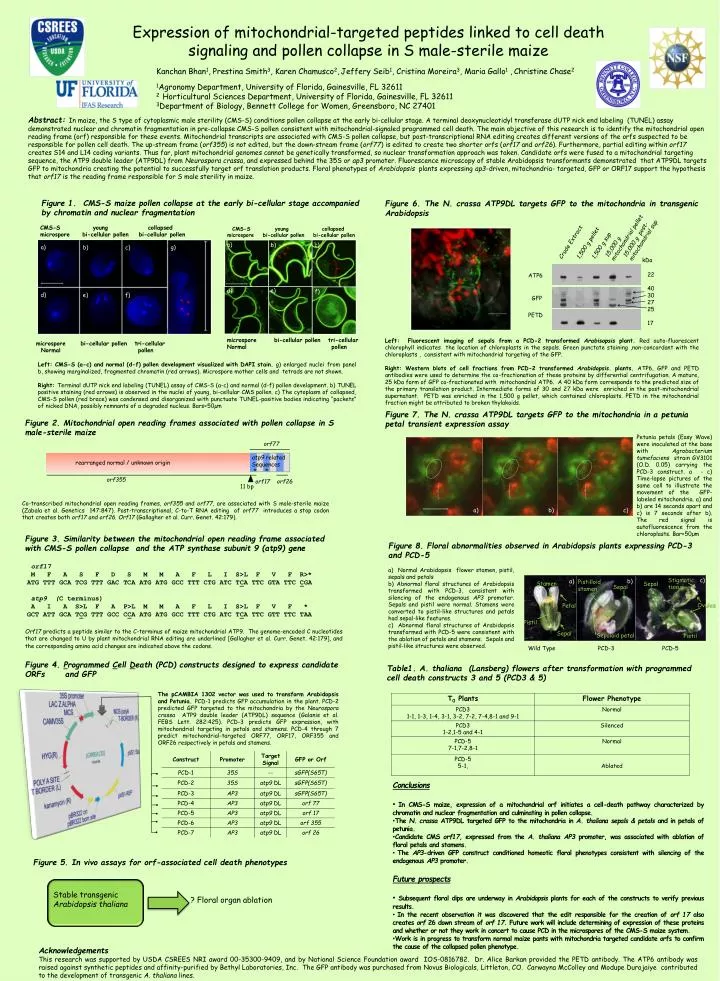
e). d). f). Expression of mitochondrial-targeted peptides linked to cell death signaling and pollen collapse in S male-sterile maize. Kanchan Bhan 1 , Prestina Smith 3 , Karen Chamusco 2 , Jeffery Seib 1 , Cristina Moreira 3 , Maria Gallo 1 , Christine Chase 2

E N D
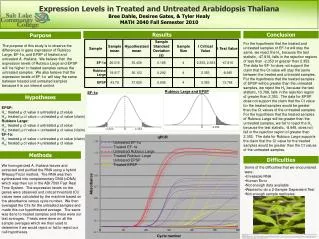
e) d) f) Expression of mitochondrial-targeted peptides linked to cell death signaling and pollen collapse in S male-sterile maize Kanchan Bhan1, Prestina Smith3, Karen Chamusco2,Jeffery Seib1, Cristina Moreira3, Maria Gallo1 ,Christine Chase2 1Agronomy Department, University of Florida, Gainesville, FL 32611 2 Horticultural Sciences Department, University of Florida, Gainesville, FL 32611 3Department of Biology, Bennett College for Women, Greensboro, NC 27401 a) b) c) Abstract: In maize, the S type of cytoplasmic male sterility (CMS-S) conditions pollen collapse at the early bi-cellular stage. A terminal deoxynucleotidyltransferasedUTP nick end labeling (TUNEL) assay demonstrated nuclear and chromatin fragmentation in pre-collapse CMS-S pollen consistent with mitochondrial-signaled programmed cell death. The main objective of this research is to identify the mitochondrial open reading frame (orf) responsible for these events. Mitochondrial transcripts are associated with CMS-S pollen collapse, but post-transcriptional RNA editing creates different versions of the orfs suspected to be responsible for pollen cell death. The up-stream frame (orf355) is not edited, but the down-stream frame (orf77) is edited to create two shorter orfs (orf17 and orf26). Furthermore, partial editing within orf17 creates S14 and L14 coding variants. Thus far, plant mitochondrial genomes cannot be genetically transformed, so nuclear transformation approach was taken. Candidate orfs were fused to a mitochondrial targeting sequence, the ATP9 double leader (ATP9DL) from Neurosporacrassa, and expressed behind the 35S or ap3 promoter. Fluorescence microscopy of stable Arabidopsis transformants demonstrated that ATP9DL targets GFP to mitochondria creating the potential to successfully target orf translation products. Floral phenotypes of Arabidopsis plants expressing ap3-driven, mitochondria- targeted, GFP or ORF17 support the hypothesis that orf17 is the reading frame responsible for S male sterility in maize. a) g) b) c) Figure 6. The N. crassa ATP9DL targets GFP to the mitochondria in transgenic Arabidopsis Figure 1. CMS-S maize pollen collapse at the early bi-cellular stage accompanied by chromatin and nuclear fragmentation CMS-S young collapsed microspore bi-cellular pollen bi-cellular pollen CMS-S young collapsed microspore bi-cellular pollen bi-cellular pollen d) e) f) 15,000 g mitochondrial pellet 15,000 g post-mitochondrial sup Crude Extract 1,500 g pellet 1,500 g sup ATP6 GFP PETD kDa microspore bi-cellular pollen tri-cellular Normal pollen microspore bi-cellular pollen tri-cellular Normal pollen 22 40 30 27 25 17 Left: Fluorescent imaging of sepals from a PCD-2 transformedArabisopsisplant. Red auto-fluorescent chlorophyll indicates the location of chloroplasts in the sepals. Green punctate staining ,non-concordant with the chloroplasts , consistent with mitochondrial targeting of the GFP. Right: Western blots of cell fractions from PCD-2 transformed Arabidopsis. plants. ATP6, GFP and PETD antibodies were used to determine the co-fractionation of these proteins by differential centrifugation. A mature, 25 kDa form of GFP co-fractionated with mitochondrial ATP6. A 40 kDa form corresponds to the predicted size of the primary translation product. Intermediate forms of 30 and 27 kDa were enriched in the post-mitochondrial supernatant. PETD was enriched in the 1,500 g pellet, which contained chloroplasts. PETD in the mitochondrial fraction might be attributed to broken thylakoids. Left:CMS-S (a-c) and normal (d-f) pollen development visualized with DAPI stain. g) enlarged nuclei from panel b, showing marginalized, fragmented chromatin (red arrows). Microspore mother cells and tetrads are not shown. Right: Terminal dUTP nick end labeling (TUNEL) assay of CMS-S (a-c) and normal (d-f) pollen development. b) TUNEL positive staining (red arrows) is observed in the nuclei of young, bi-cellular CMS pollen. c) The cytoplasm of collapsed, CMS-S pollen (red brace) was condensed and disorganized with punctuate TUNEL-positive bodies indicating “packets” of nicked DNA, possibly remnants of a degraded nucleus. Bars=50µm Figure 7. The N. crassa ATP9DL targets GFP to the mitochondria in a petunia petal transient expression assay Figure 2. Mitochondrial open reading frames associated with pollen collapse in S male-sterile maize Petunia petals (Easy Wave) were inoculated at the base with Agrobacteriumtumefaciensstrain GV3101 (O.D. 0.05) carrying the PCD-3 construct. a - c) Time-lapse pictures of the same cell to illustrate the movement of the GFP-labeled mitochondria. a) and b) are 14 seconds apart and c) is 7 seconds after b). The red signal is autofluorescence from the chloroplasts. Bar=50µm orf77 • atp9 related • Sequences • * * rearranged normal / unknown origin orf355 orf17 orf26 11 bp Co-transcribed mitochondrial open reading frames, orf355 and orf77, are associated with S male-sterile maize (Zabala et al. Genetics 147:847). Post-transcriptional, C-to-T RNA editing of orf77 introduces a stop codon that creates both orf17 and orf26. Orf17 (Gallagher et al. Curr. Genet. 42:179). a) b) c) Figure 8. Floral abnormalities observed in Arabidopsis plants expressing PCD-3 and PCD-5 Figure 3. Similarity between the mitochondrial open reading frame associated with CMS-S pollen collapse and the ATP synthase subunit 9 (atp9) gene Orf17 predicts a peptide similar to the C-terminus of maize mitochondrial ATP9. The genome-encoded C nucleotides that are changed to U by plant mitochondrial RNA editing are underlined [Gallagher et al. Curr. Genet. 42:179], and the corresponding amino acid changes are indicated above the codons. Stigmatic tissue c) b) a) Pistilloid stamen Stamen Sepal Sepal orf17 M F A S F D S M M A F L I S>L F V F R>* ATG TTT GCA TCG TTT GAC TCA ATG ATG GCC TTT CTG ATC TCA TTC GTA TTC CGA atp9 (C terminus) A I A S>L F A P>L M M A F L I S>L F V F * GCT ATT GCA TCG TTT GCC CCA ATG ATG GCC TTT CTG ATC TCA TTC GTT TTC TAA a) Normal Arabidopsis flower stamen, pistil, sepals and petals b) Abnormal floral structures of Arabidopsistransformed with PCD-3, consistent with silencing of the endogenous AP3 promoter. Sepals and pistil were normal. Stamens were converted to pistil-like structures and petals had sepal-like features. c) Abnormal floral structures of Arabidopsis transformed with PCD-5 were consistent with the ablation of petals and stamens. Sepals and pistil-like structures were observed. Ovules Petal Pistil Pistil Sepal Sepaloid petal Pistil Wild Type PCD-3 PCD-5 Figure 4. Programmed Cell Death (PCD) constructs designed to express candidate ORFs and GFP Table1. A. thaliana (Lansberg) flowers after transformation with programmed cell death constructs 3 and 5 (PCD3 & 5) The pCAMBIA 1302 vector was used to transform Arabidopsis and Petunia. PCD-1 predicts GFP accumulation in the plant. PCD-2 predicted GFP targeted to the mitochondria by the Neurosporacrassa ATP9 double leader (ATP9DL) sequence (Galanis et al. FEBS Lett. 282:425). PCD-3 predicts GFP expression, with mitochondrial targeting in petals and stamens. PCD-4 through 7 predict mitochondrial-targeted ORF77, ORF17, ORF355 and ORF26 respectively in petals and stamens. • Conclusions • In CMS-S maize, expression of a mitochondrial orf initiates a cell-death pathway characterized by chromatin and nuclear fragmentation and culminating in pollen collapse. • The N. crassaATP9DL targeted GFP to the mitochondria in A. thaliana sepals & petals and in petals of petunia. • Candidate CMS orf17, expressed from the A. thaliana AP3 promoter, was associated with ablation of floral petals and stamens. • The AP3-driven GFP construct conditioned homeotic floral phenotypes consistent with silencing of the endogenous AP3 promoter. • Future prospects • Subsequent floral dips are underway in Arabidopsis plants for each of the constructs to verify previous results. • In the recent observation it was discovered that the edit responsible for the creation of orf 17 also creates orf 26 down stream of orf 17. Future work will include determining of expression of these proteins and whether or not they work in concert to cause PCD in the microspores of the CMS-S maize system. • Work is in progress to transform normal maize pants with mitochondria targeted candidate orfs to confirm the cause of the collapsed pollen phenotype. Figure 5. In vivo assays for orf-associated cell death phenotypes Stable transgenic Arabidopsis thaliana ? Floral organ ablation Acknowledgements This research was supported by USDA CSREES NRI award 00-35300-9409, and by National Science Foundation award IOS-0816782. Dr. Alice Barkan provided the PETD antibody. The ATP6 antibody was raised against synthetic peptides and affinity-purified by Bethyl Laboratories, Inc. The GFP antibody was purchased from Novus Biologicals, Littleton, CO. CarwaynaMcColley and ModupeDurojaiye contributed to the development of transgenic A. thaliana lines.