Download

1 / 51

E N D
T-cell Receptor Feb 9, 2004
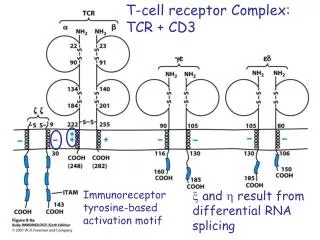
T-cell Receptor • The biochemical signals that are triggered in T cells by antigen recognition are transduced not by the T cell receptor itself but by invariant proteins called CD3 an z (zeta), which are noncovalenlty linked to the antigen receptor to form the TCR complex.
TCR • Mature T cells express one of two types of TCR: a heterodimer composed either of a and b chains or of g and d chains. • Because T cells expressing ab receptors account for 90% of T-cell helper function and cytotoxic activity, the major focus of this discussion will be on this type of TCR. • The gd T cells, whose physiologic role is still unclear, will be reviewed later on.
T-cell Receptor • T cells also express other membrane receptors that do not recognize antigen but participate in responses to antigens: these are collectively called accessory molecules.
T-cell • Therefore, we will first focus on the TCR followed by a discussion on accessory molecules
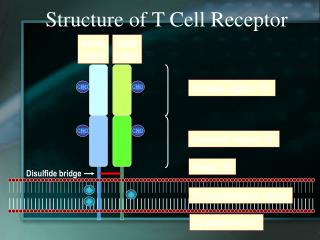
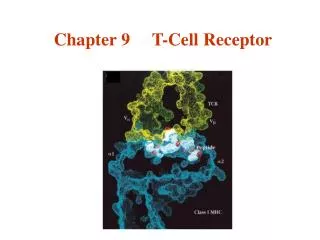
TCR • The antigen receptor of MHC-restricted CD4+ helper T cells and CD8+ cytotoxic lymphocytes is a heterodimer • As mentioned before the TCR consists of two transmembrane polypeptide chains, designated a and b, covalently linked to each other by disulfide bonds.
TCR • Each a and b chain consists of one ‘Ig-like’ N-terminal variable region (V), one Ig-like constant (C) domain, a hydrophobic transmembrane region, and a short cytoplasmic region. • Thus the extracellular portion of the ab heterodimer is structurally similar to the antigen-binding fragment (Fab) of an Ig, which is made up of the V and C regions of a light chain and the V region and one C region of a heavy chain.
TCR • The V region of the TCR a and b chains contain short stretches of amino acids where the variability between different TCRs is concentrated, and these form the hypervariable or complementarity-determining regions (CDRs). • Three CDRs in the a chain are juxtaposed to three similar regions in the b chain to form the peptide recognizing complex.
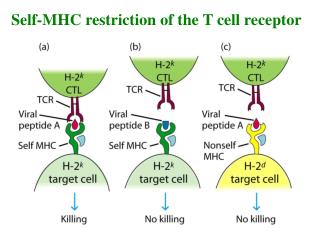
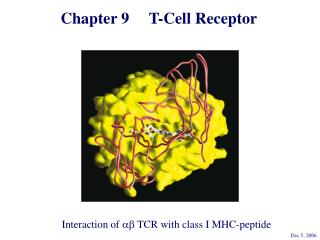
TCR • An analysis of TCR sequence diversity has shown that the vast majority of amino acid variation resides in the region between the V- and J-region gene segments, which corresponds to the CDR3 regions of antibodies. • This has led to models in which the CDR3 loops of Va and Vb make the principal contacts with the antigenic peptide bound to the MHC
TCR-MHC Interactions • The CDR3 loops of Va and Vb make the principal contacts with the antigenic peptide bound to the MHC.
TCR α & β GENES & THE GENERATION OF TCR DIVERSITY • To generate the diversity of TCRs required to recognize a wide spectrum of antigenic determinants, the TCRa and b genes use a strategy of recombination similar to that of the immunoglobulin genes. • The germline TCRb-gene locus contains 20-30 V (variable), 2 D (diversity), and 13 J (joining) gene segments
Rearrangement of the TCR α and β genes. • The TCR a-gene locus contains multiple V and J segments, only several of which are shown here. Similarly, the TCR b-gene locus contains multiple V, D, and J segments. • During T-cell ontogeny, the TCR genes rearrange (arrows), so that one of the Va segments pairs with the Ja segment and a Vb segment pairs with a Db and Jb segment. The two C (constant) segments in the b gene are very similar, and differential use of Cb1 and Cb2 does not contribute to TCR diversity.
CD3 • TCRs occur as either of two distinct heterodimers, ab or gd, both of which are expressed with the non-polymorphic CD3 polypeptides g, d, e, and z. • The CD3 polypeptides, especially z and its variants, are critical for intracellular signaling.
T-cell Receptor
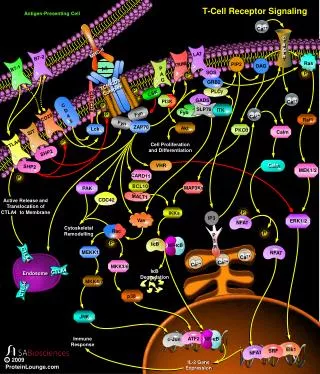
SIGNAL TRANSDUCTION BY THE TCR • Key to the ability of the TCR to deliver intracellular signals is its interactions with protein tyrosine kinases (PTKs). • In unstimulated T cells, Fyn, a member of the Src family of PTKs, associates with the cytoplasmic domains of CD3 chains. • A second Src-like PTK, called Lck, binds to the cytoplasmic domains of CD4 and CD8 and thus can be brought into proximity with the TCR through the interactions of these coreceptors with the MHC.
SIGNAL TRANSDUCTION BY THE TCR • Stimulation of the TCR by antigen-MHC triggers the phosphorylation of tyrosine residues in the cytoplasmic domains of the CD3 chains of the receptor complex. • According to a widely accepted model of TCR signaling, Lck and Fyn are responsible for these initial phosphorylation events.
gd TCR • The gd TCR are a second type of TCR. • Their function remains largely unresolved. • They do not recognize MHC-associated peptides and are not MHC restricted. • In mice and chickens they are found in the small bowel mucosa and termed intraepithelial lymphocytes. • In humans they are found in the tongue, uterus and vagina.
gd TCR • In mice many gd TCR T-cells develop in neonatal life and express one particular TCR with essentially no variability in the V region. • Therefore it is not known whether these subsets perform different T-cell function.
Accessory Molecules
CD45 • CD45 is a large (180-220 kd) transmembrane cell surface molecule that is expressed by all leukocytes, including all T lymphocytes. • The cytoplasmic domain of CD45 has tyrosine phosphatase activity. • CD45 activity is at the very early steps of TCR signaling, indicating that CD45 is required for the functional coupling of the TCR and its PTKs.
CD45 • Memory and naive T cells also differ in their surface phenotypes, most notably in their expression of CD45 isoforms. • Alternative splicing of CD45 mRNA gives rise to a number of different isoforms of CD45 that differ in the size and composition of their extracellular domains. • Naive T cells express 205- to 220-kd isoforms designated CD45RA, whereas memory T cells express a 180-kd isoform called CD45RO.
COSTIMULATION BY CD28 • Despite their complexity, the signals delivered by the TCR are insufficient to fully activate T cells. • Rather, T-cell activation requires the delivery of both the TCR signals and a second set of signals generated by costimulatory molecules. • In the absence of the proper costimulus, stimulation of the TCR alone can induce a T cell to enter a state in which it remains viable but is refractory to stimulation by antigen. This state, which is known as anergy, can be long-lived, persisting for weeks to months in vitro.
COSTIMULATION BY CD28 • The best characterized (and probably the most important) costimulatory molecule is CD28, a 44-kd glycoprotein that is expressed as a homodimer on the surfaces of virtually all CD4 T cells and approximately 50% of CD8 T cells. • CD28 binds two distinct cell surface molecules, B7.1 and B7.2, found on dendritic cells, macrophages, and activated B cells. • The combination of TCR stimulation and the interaction of CD28 with its B7 ligands fully activates T cells and results in substantially greater lymphokine production than can be induced by TCR signals alone
CTLA-4 • The number of antigen-specific T cells falls dramatically when an immune response terminates. • Following successful clearance of virus, the number of virus-specific CTLs in a mouse can drop from 108 to 106—a decrease of 99%. • The decline reflects apoptosis, perhaps triggered by cytokine withdrawal or by engagement of Fas or other members of the tumor necrosis factor (TNF) receptor family.
CTLA-4 • One important negative regulator of T-cell activation is, a T-cell surface molecule induced on activation and not found on resting cells. • CTLA-4 shares considerable sequence homology with CD28 and, like CD28, binds B7.1 and B7.2 on the APC. • Unlike CD28, however, CTLA-4 delivers inhibitory signals to T cells, so that engagement of CTLA-4 tends to strongly diminish T-cell responses. • Mice genetically engineered to lack CTLA-4 die with massive polyclonal expansion of T lymphoblasts.
CD2 • CD2 is a glycoprotien present on more than 90% of mature T-cells and 50-70% of thymocytes. • This molecule contains two extracellular Ig domains. • The principle ligand for CD2 is LFA-3 (CD58).
CD2 • CD2 functions both as an adhesion molecule and signal transducer. • The association of CD2 with the TCR complex helps to aggregate the TCR in the regions of cell–cell contact, allowing the stabilization of low-affinity TCR/MHC interactions. • Finally, CD2 is involved in the regulation of cytokine production by T cells. • Stimulation via the CD2 pathway can skew the cytokine profile toward a TH2-like phenotype.
Integrins • We have already discussed integrins in the context of neutrophils. • The major functions of T-cell integrins are to mediate adhesion to APCs, endothelial cells, and extracellular matrix proteins. • The avidity of integrins for their ligands is increased rapidly on exposure of the T-cells to cytokines called chemokines and after stimulation of T-cells through the TCR.
Integrins Figure 6-11
Integrins • Integrins will be discussed on Wednesday in more detail.
CD44 • CD44 is expressed by activated and memory cells in comparison to naïve cells. • This molecule is responsible for retension of T cells in extravascular tissues at sites of infection and for the binding to endothelial cells at sites of infection and in mucosal tissues.
CD40L • The CD40L on T-cells binds to the CD40 on B-cells thus an important mediator of stimulation of B cells. • We have covered the CD40L related to our PBL.
CD95 (Fas receptor) • Activated T cells also express a ligand for death receptor Fas (CD95). Engagement of Fas by Fas ligand on T-cells results in apoptosis and is important for eliminating T-cells. • FasL also provides one o the mechanisms by which CTLs kill targets.
T-cell Subtypes • T helper Th1 cells secrete pro-inflammatory cytokines (IFN-g, TNF, and IL-2. • Whereas T helper Th2 cells produce cytokines that generally stimulate Ig responses (IL-4, -5, -6, -9, and -10). • These biases tend to be self-reinforcing: IL-10 represses Th1 cell activity and IFN-g inhibits Th2 cells.
T-cells • It is not clear whether the Th1/Th2 distinction corresponds to a simple dichotomy or rather to two extreme poles, between which intermediate patterns of cytokine production can be found. • In addition, there is mounting evidence for other helper classes.