Download

1 / 48
480 likes | 565 Views
Microbial Genomics and Secondary Metabolites MedILS, Split, Croatia June 29, 2007. First insights into bacterial Ser/Thr/Tyr phosphoproteome. Boris Ma č ek Department of Proteomics and Signal Transduction Max Planck Institute of Biochemistry Martinsried, Germany. Our workflow: „GeLC-MS“.

E N D
Microbial Genomics and Secondary Metabolites MedILS, Split, Croatia June 29, 2007 First insights into bacterial Ser/Thr/Tyr phosphoproteome Boris Maček Department of Proteomics and Signal Transduction Max Planck Institute of Biochemistry Martinsried, Germany
Our workflow: „GeLC-MS“ Aebersold R, Mann M.2003.Nature 422: 198-207
High-resolution, accurate, fast scanning MS: FT-MS Hybrid linear ion trap FT-MS instruments Electrostatic field: Electromagnetic field: LTQ-FTICR MS Olsen JV et al., MCP2005 Olsen JV et al., MCP2004 Non-destructive Detection: Parts per million mass accuracy In a 7-Tesla magnetic field an ion with m/z =100 will spin1,000,000 cycles (travel ~30 km) in a1 sec. observation period
High-mass accuracy – why is it important? Consider all theoretical tryptic peptide masses from the human IPI database (> 40,000 protein sequence entries) Example: Tryptic HSP-70 peptide: ELEEIVQPIISK, mass 1396.7813 Da
Quantitation with Stable Isotope Labeling Unlabeled peptide: Labeled peptide:
Stable Isotope Labeling byAmino Acids in Cell Culture (SILAC) Stable isotope dilution: same physico-chemical properties Upregulated protein. Peptide ratio >1 Arg- 13C6 ”normal AA” ”heavy AA” Arg-12C6 Arg-13C6 Resting cells Treated (drug, GF) Arg- 12C6 Combine and lyse, protein purification or fractionation Background protein. Peptide ratio 1:1 Arg- 12C6 Arg- 13C6 Proteolysis (trypsin, Lys-C, etc.) Quantitation and identification by MS (nanoscale LC-MS/MS) Ong SE et. al., Mol Cell Proteomics 2002 m/z
SILAC requirements • Cell/organism must be auxotrophic for the corresponding AA • Growth in defined media lacking the SILAC labeling amino acid (e.g. Arg, Lys) • Stable Isotope Labeled Amino Acids: • Growth supplements (e.g. dialyzed serum) if necessary Arg-13C6 (Δm=6 Da) Arg-13C615N4 (Δm=10 Da) Lys-13C6 (Δm=6 Da) Lys-13C615N2 (Δm=8 Da)
Gel-free phosphoproteome analysis workflow Cell harvest & trypsin digestion Strong cation exchange Chromatography pH<3 TiO2 Chromatography pH<3 (bind) pH>10 (elute) Cell culture Data Analysis LC-MS pH~1 ½ day 1-2 days ½ day days ½ - 1 day O.N. 1-2 days
Phosphopetide enrichment by Titansphere (TiO2) chromatography Competitive binding of peptides with DHB < < Larsen et al. (2005) Mol Cell Proteomics 4:873-886
LC separation • Proxeon nano-ESI source • Agilent 1100, Proxeon nano-HPLC systems • self-packed 75 μm x ~10 cm Porous C18 HPLC columns • flow ~250 nL/min
Hybrid linear ion trap FTICR MS: LTQ FT (Thermo Scientific)
LTQ-FT MS/MS optimized scan cycle: FT-MS MS-Full SIM-MS 1st SIM-MS 2nd SIM-MS 3rd IT-MS MS2 MS2 MS2 0 300 600 900 1200 1500 1800 Time [msec] LTQ-FT data-dependent experiments Two Mass Spectrometers in one - High duty-cycle Ion trap MS: + sensitivity (MS/MS mode) and speed resolution, mass accuracy and dynamic range FTICR MS: + resolution, mass accuracy and dynamic range sensitivity (MS/MS mode) and speed LTQ-FT: The best from both instruments
Beausoleil SA et al. (2004) PNAS 101:12130-35. Phosphopeptide-directed MS3
Recent advances in FT-MS: LTQ-Orbitrap (Thermo) Non-destructive Detection: Orbitrap: FT: FT: Full scan Full Full Full Full SIM1 SIM1 SIM1 SIM1 SIM2 SIM2 SIM2 SIM2 SIM3 SIM3 SIM3 SIM3 LTQ: LTQ: LTQ: LTQ: 2 2 2 2 MS MS MS MS MS MS MS MS2 MS MS MS2 MS MS MS MS MS2 MS2 MS MS MS 2 2 2 2 3 3 3 3 2 2 2 2 3 3 3 3 2 2 2 2 3 3 3 3 MS MS MS MS MS MS MS MS MS MS MS MS MS MS MS MS MS MS MS MS MS MS MS MS 0 1 2 0 1 2 0 1 2 0 1 2 Time [s] Time [s] Time [s] Time [s]
LTQ-Orbitrap in the analysis of PTMs „Hot“ CID Multi-stage activation
30ms 30ms 30ms 30ms wb m/z Precursor - 32.6 Da - 49 Da - 98 Da CID with Multi-Stage Activation (MSA) Pseudo MS3 Easy to identify multiply-phosphorylated peptides: 4, 5 and 6 phosphates TiO2-enrichment of flow through from SCX (HeLa_EGF_CE_0_5_10)
CID in the C-trap (”Hot” CID or HCD) Informative low mass ions – reporter ions (Phosphotyrosine immonium ion, m/z = 216.0426)
Intracellular signaling networks (EGFR, HeLa) (www.phosida.com) • identified more than 2200 phosphoproteins • determined more than 6600 phosphorylation sites • pS (87%)/pT (12%)/pY (1.5%) • less than 15% sites regulated by EGF treatment • → systems biology modeling of signaling networks Olsen et al. (2006) Cell 127(3):635-648
Protein phosphorylation in bacteria Two-component system
Protein phosphorylation in bacteria Phosphoenolpyruvate:carbohydrate phosphotransferase system (PTS)
Overview of Ser/Thr/Tyr phosphorylation in prokaryotes • many putative Ser/Thr/Tyr kinases identified (mostly in silico) • 2D gel studies suggest presence of hundred(s) of phosphoproteins • However: • only about 150 proteins from about 35 species shown to be phosphorylated • only about 70 Ser/Thr/Tyr phosphorylation sites identified • phosphorylation analysis mostly in vitro! • → clear need for in-depth detection and characterization of protein • phosphorylation in vivo
Ser/Thr/Tyr phosphorylation in B. subtilis *Macek et al. 2006. Mol Cell Proteomics 6(4): 697-707
Ser/Thr/Tyr phosphorylation in B. subtilis *Macek et al. 2006. Mol Cell Proteomics 6(4): 697-707
y*19 y*18 y*17 y14 y13 y11 y7 y6 y5 y4 y3 y2 V T A D pS G I H A R P A T V L V Q T A S K y14++ 740.427 100 90 80 y*18++ y*19+++ y*18+++ 70 917.504 635.683 y13++ y*17+++ 60 y7 573.662 Relative Abundance 50 y16++ y2 825.480 40 y4 234.145 y11 y14+++ 406.229 y5 30 1114.647 y*17++ y3 20 305.182 946.560 10 1017.598 0 200 300 400 500 600 700 800 900 1000 1100 m/z Hpr protein Orbitrap full scan C-trap MS/MS(HCD) Precursor m=0.91ppm Fragment m<2ppm
y7 y6 y5 y3 y8 pS V I V N A L R K b2 b3 b4 b6 b8 CodY – Global regulator of transcription FT-ICR full scan ion-trap MS/MS (CID) Precursor m=6.39 ppm Fragment m<0.5 Da
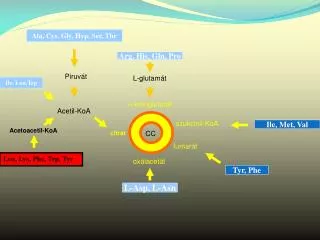
Phosphorylationin the main pathways of carbohydrate metabolism (B. subtilis) GLYCOLYSIS Enolase (eno) L-lactate dehydrogenase (lctE) Triose phosphate isomerase (tpi) G-3-P dehydrogenase (gap) Pyruvate kinase (pykA) Malate dehydrogenase (citH) Phosphoglycerate mutase (pgm) Glucose-6-phosphate isomerase (pgi) Fructose-bisphosphate aldolase (fbaA) Pyruvate dehydrogenase (pdhB) Phosphoglycerate kinase (pgk) Phosphoglucomutase (ybbT) TCA CYCLE Citrate synthase II (citZ) Succinyl-CoA synthetase (sucC, sucD)
E. coli vs. B. Subtilis phosphoproteome • phosphoproteomes similar in: • size • distribution of S/T/Y phosphorylation • classes of phosphorylated proteins • increased essentiality *Macek et al. 2007. submitted
Evolutionary conservation of bacterial S/T/Y phosphoproteins • test set of 9 archaeal, 53 bacterial and 8 eukaryotic proteomes • look for orthologs of bacterial phosphoproteins (2-directional BLAST; Needle) • reported as average % of identified phosphoprotein orthologs in tested species • compared to the random protein population E. coli phosphoproteome B. subtilis phosphoproteome
Evolutionary conservation of bacterial S/T/Y phosphorylation sites → phosphoserine:
Evolutionary conservation of bacterial S/T/Y phosphorylation sites → phosphothreonine:
Bacterial S/T/Y phosphoproteins with P-sites conserved from Archaea to H. sapiens • cysteinyl t-RNA synthetase • phosphoglucomutase • nucleoside diphosphate kinase • pyruvate kinase • enolase • predicted GTP-binding protein • D-3 phosphoglycerate dehydrogenase • phosphoglucosamine mutase • elongation factor Ef-Tu
→ mutases are good internal standards for “quality control”!
”normal AA” ”heavy AA” (+8Da) Lys-12C614N2 Lys-13C615N2 Treated cells (succinate or low P) Control cells Combine and lyse GeLC-MS Proteolysis (trypsin) Strong cation exchange chromatography(SCX) Titanium oxide chromatography nanoLC-MS/MS (Quantitation and identification by MS) Stable Isotope Labeling byAmino Acids in Cell Culture (SILAC): Bacillus subtilis (Arg-, Lys-) Peptide ratio 1:1 - No change. m/z Peptide ratio >1 - Downregulation. control treated
Dynamics of protein expression in B. subtilis : Growth on succinate
Dynamics of protein phosphorylation in B. subtilis : Growth on succinate
NH2 COOH H15 S46 S12 Growth on low succinate: Hpr protein Ser46: pSIMGVMSLGIAK GAEITISASGADENDALNALEETMK Ser12: VTADpSGIHARPATVLVQTASK GAEITISASGADENDALNALEETMK
Dynamics of protein expression in B. subtilis : Growth under low PO43-
Dynamics of protein phosphorylation in B. subtilis : Growth under low PO43-
NH2 COOH H15 S46 S12 Growth on low PO43-: Hpr protein Ser46: pSIMGVMSLGIAK YDADVNLEYNGK Ser12: VTADpSGIHARPATVLVQTASK YDADVNLEYNGK
Conclusions • SCX + TiO2 + FT MS - a powerful and generic strategy for phosphopeptide • enrichment and detection • bacteria posess an elaborate Ser/Thr/Tyr phosphoproteome • majority of enzymes in the main pathways of carbohydrate metabolism are • phosphorylated • enzymes of the PTS system are phosphorylated on Ser/Thr/Tyr • → possible cross-talk • Ser/Thr/Tyr phosphorylation is dynamic process • → likely regulatory role • phosphoroteins and phosphorylation sites show increased evolutionary • conservation • at least 9 P-sites conserved from Archaea to man: ancient regulatory role?
Acknowledgements Max-Planck-Institute for Biochemistry Matthias Mann Florian Gnad Jesper V. Olsen Chanchal Kumar Technical University of Denmark Ivan Mijakovic Boumediene Soufi Dina Petranovic Thermo Scientific Stevan Horning Oliver Lange