Download
1 / 47
540 likes | 1.18k Views
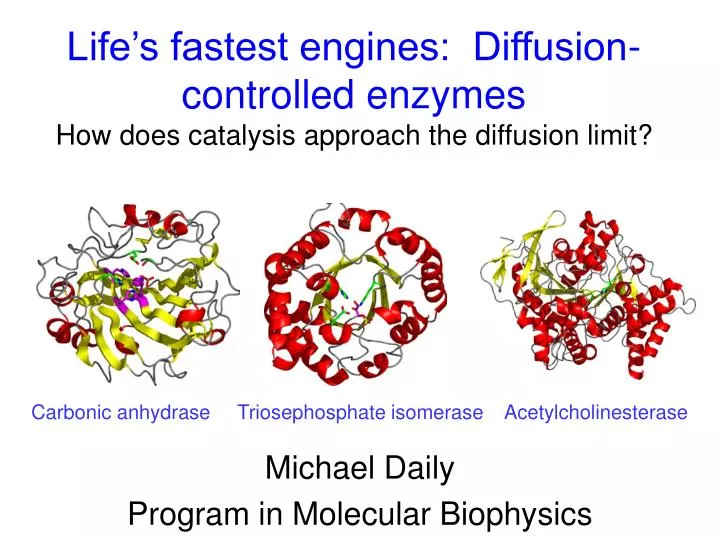
Life’s fastest engines: Diffusion-controlled enzymes How does catalysis approach the diffusion limit?. Michael Daily Program in Molecular Biophysics. Carbonic anhydrase Triosephosphate isomerase Acetylcholinesterase. Enzyme power: remarkable rate enhancement.

E N D
Life’s fastest engines: Diffusion-controlled enzymesHow does catalysis approach the diffusion limit? Michael Daily Program in Molecular Biophysics Carbonic anhydrase Triosephosphate isomerase Acetylcholinesterase
Enzyme power: remarkable rate enhancement • Some biological reactions are “geologically slow” (hundreds – 1 billion years half time) • All catalyzed reactions operate at millions per second • Transition state affinities as high as 1024 M Wolfenden & Snider (2001). Acc. Chem. Res. 34, 938-45 Ornithine decarboxylase: t1/2 = 78 million years at 25 C. Crystal structure: Almrud, J.J. et. al. (2000). JMB295, 7.
Some major enzyme problems • Bond rearrangement • Stabilization of high-energy intermediates, transition state • Orientation of reactive groups • Proton shuttling at neutral pH
Enzyme catalysis: mathematics E + Sts knon KA‡ ES‡ E+ S kcat E + P KM ES kcat/KM = knon*KA‡
Enzymes exert remarkable control over kcat/KMrelative to the variation of knon kcat/KM: 102.5 variation knon: 1014 variation Radzicka, A. and Wolfenden, G. (1995). Science267 (5194), 90-93. Diffusion-controlled enzymes
What is the limit of the molecular encounter rate? • According to Smolouchowski equation, E-S collision rate is limited to ~109/s 1 2) Orientational constraints limit the reactive encounter rate to ~106/s 2 3) Electrostatic attraction or guidance of S to E can raise the diffusion limit to 108-109/s. 3 Alberty, R.A. and Hammes, G.G. (1958). J. Phys Chem62, 154-59
Electrostatics cause large differences in barnase-barstar association rates Wild type (top) and barnase mutants ~1000x variation in k1 at 0.1M ionic strength Basal k1 ~ 105 M-1s-1 Schreiber, G. and Fersht, A.R. (1996). Nat. Struct. Biol.3 (5), 427-431.
Summary: enzymes overcome two physical problems • Chemical efficiency • Geometry (entropic problem) • High-energy intermediates (enthalpic problem) • Enzyme rate does not appear to be limited by knon • Molecular encounter • Coulomb’s law (enthalpic effect) • Steering (entropic effect)
Carbonic Anhydrase II – an introductory case • CO2 control – very important metabolically • Ubiquitous, independently evolved 3 times • Very fast uncatalyzed reaction (t1/2 = 5s) • Rapid, efficient proton shuttling Crystal structure: Hakansson, K.et. al. (1992). J. Mol. Biol. 227, 1192.
The CA II reaction has two major steps Silverman, D.N. and Lindskog,S. (1988). Acc. Chem. Res. 21, 30-36. Tu, C. et al. (1989). Biochemistry28, 7913-18. CO2 hydration CO2 + OH- <-> HCO3- Rate-limiting nucleophile regeneration Zn-H2O <-> Zn-OH + H+
Zinc hydroxide nucleophile: A well-understood problem T199 orients Zn-bound OH- to attack CO2 T199A mutant has ~0.01x the kcat/KM of wt E106 anchors T199 E106A,E106Q mutants have ~0.1x the kcat/KM of wt Catalytic residues (E106,T199,H64) Zn ion and ligands Xue et. al. (1993). Proteins17 (1), 93-106. Krebs et. al. (1993). J. Biol. Chem. 268 (36), 27458-66. Liang et. al (1993). Eur. J. Biochem. 211 (3), 821-7. 2bcc.pdb
Nucleophile regeneration: problems and solutions • Proton transfer from Zn(OH2) to solution OH- is limited to 103/s • Concentrated solution buffers deprotonate Zn(OH2) at as high as 106/s (Jonsson et. al 1976) • Solution buffer cannot efficiently penetrate to the buried Zn(OH2) active center • Active site waters and H64 efficiently shuttle protons from the active center to the surface (Tu et al. 1989, Silverman 1995, Jackman et. al 1996, Skolnick et. al 1996).
His 64-water network nucleophile regeneration mechanism Catalytic residues Proton shuttle H64 Bulk solvent Four water molecules transfer protons from catalytic site to his 64 Catalytic water
Unsolved problems of CA II • CO2 hydration- • Position of CO2 (weak binding) • Proton transfer • Effect of moving the proton shuttle • Which waters are involved in proton transfer? • Quantum mechanical mechanistic details • Beyond structural biology
CA II is a well-understood enzyme (from a structural biology viewpoint) • Catalytic groups identified • Function, importance of catalytic groups known • Exact details of proton transfer still being researched • Quantum mechanical mechanism still being researched • Mechanism is probably understood at the design level
Triosephosphate Isomerase (TIM)An intermediate case • Ubiquitous (glycolytic enzyme) • Large rate enhancement (~1013) • Two difficult proton transfers • Paradoxical E-S electrostatic attraction Catalytic residues (H95, E165, K12) Substrate (DHAP)
TIM mechanism involves two difficult proton transfers C1 deprotonation (rate limiting) DHAP enediolate DHAP O1-O2 proton transfer GAP GAP Knowles, J.R. (1991). Nature 350, 121-124.
TIM catalysis solves three problems • Difficult C1 deprotonation (pKa ~ 20) • Unstable negatively charged enediolate intermediate • Proton transfer from O1 to O2 (pKa ~ 14)
The crucial C1 deprotonation step is well-understood H95 and K12 polarize C2 carbonyl, lowering C1 pKa from ~20 to ~14 Unusually close E165-C1 contact prepares for C1 deprotonation 1.2A Crystal structure: Jogl et. al (2003). PNAS100 (1), 50-55 Nickbarg et al (1988), Lodi & Knowles (1991)
A multi-pronged positive field stabilizes negative enediolate positive end of helix dipole K12+ stabilizes O2- O1, O2 hbond with H95
O1-O2 proton transfer: two pathways Low-barrier hbond (LBHB): between H950 and enediolate (both pKa ~14) facilitates H95-O2 proton transfer Neutral H95 moves proton from O1 (OH) to O2- E165 is known to participate in O1-O2 transfer some fraction of the time. Harris et. al. (1998), Cui and Karplus (2001).
Electrostatics: paradox of E-S attraction in TIM Yeast TIM has net charge of -6 at pH 7. Oxygen atoms Nitrogen atoms Substrate (DHAP) 1NEY.pdb (yeast TIM)
TIM-GAP attraction is dominated by potential in active site vicinity • TIMs of varying charge (-12 to +12) have rate enhancements from 100-1000 • Attractive field is calculated near the active site and within 10A of the enzyme • Biological application: TIM can function at diffusion limit in different pH environments (Wade et. al 1998, Proteins31:406-416)
TIM: answered questions • Catalysts: E165 and H65 • Rate-limiting step: C=O polarization stimulates α-C deprotonation • Enediolate stabilization: multi-pronged positive field • E-S encounter mechanism – Active site region is crucial in attracting S
TIM: unresolved problems • O1-O2 proton transfer • E165 and H95 are both involved, but at what ratio: probably a random element • Role of low-barrier H-bond (LBHB) • Does this explain how a neutral H95 can protonate the enediolate? • Highly controversial concept
Acetylcholinesterase – diffusion control and amazing catalytic power • Remarkable catalytic proficiency (~1017) • Difficult nucleophilic step, unstable intermediate • Electrostatic E-S attraction and guidance Harel et. al. (2000) Catalytic triad (S200, H440, E327)
Ache catalytic mechanism: nucleophilic carbonyl hydrolysis choline acetylcholine acetate acetyl- choline Oxyanion 1 acetate choline Sussman et. al (1991). Science253, 872-879. Oxyanion 2
Ache facilitates a difficult hydrolysis and stabilizes buried substrate charges • S200 nucleophile polarization and deprotonation: the rate-limiting step • Oxyanion intermediate stabilization: a negative charge in a buried active site • Quaternary ammonium stabilization: a positive charge in a buried active site
Ache catalytic triad facilitates a difficult nucleophilic attack S200: the nucleophile Transition state analog: TMTFA E327: short, strong H-bond stabilizes H440+ (Massiah et. al 2001) H440 polarizes and deprotonates S200-OH nucleophile Crystal structure: Harel et. al (1996). JACS118, 2340-6.
Ache oxyanion hole stabilizes high-energy tetrahedral intermediate acetylcholine oxyanion Mechanism could be H-bond, dipole, or concerted proton transfer to O- Ordentlich et. al (1998). JBC273 (31), 19509-17
Ache catalytic center:A serine protease … sort of Mirror image catalytic triads Chymotrypsin AChE Two-pronged oxyanion hole Three-pronged oxyanion hole Pdb: Yennawar et. al (1994). Ref: Sussman et. al 1991. Pdb: Harel et. al (2000).
TMA binding pocket: dealing with a buried positive charge H-bonds to waters Cation-π interactions Coulombic interaction S200 nucleophile Harel et. Al (1993). PNAS90(19),9031-5.
E-S attraction: Coulombic attraction and aromatic guidance Negative charges on surface (red) attract positive acetylcholine Aromatic residues in gorge make cation-π interactions with acetylcholine
Electrostatic potential gradient guides acetylcholine into the active site Figure 8 Felder et al (1997). J. Mol. Graph. Model. 15, 318-327. Also see Radic et. al (1997), Ripoll et. al (1993).
Acetylcholinesterase is a well-understood enzyme • Catalytic triad facilitates difficult proton transfers • Oxyanion stabilization • TMA binding • E-S attraction: Coulomb’s law and guidance (Felder).
Some “minor details” are still being worked out • How much do short, strong hydrogen bonds (SSHBs) in the active site facilitate proton transfer? • What is the precise mechanism of the oxyanion hole? • What exact matrix of interactions gives rise to acetylcholine – AChE attraction?
Superoxide dismutase – designed to be super-perfect • Converts O2- to H2O2, Cu and Zn-dependent • Asymmetric potential distribution -> E- attracts S- (Getzoff et. Al 1983, Klapper et. Al 1986) • By reducing negative charge near active center, ke can be raised to 2*109s-1. (Getzoff et. Al 1992, Nature358, 347-351). Structure: Tainer et. al. (1982). JMB 160, 181.
Enzyme catalytic mechanisms: qualitatively understood, quantitatively imprecise • Proton transfers – pKa lowering, proton shuttling, networking, low-barrier H-bond • Bond polarization – activate nucleophile, increase electrophilicity, lower pKa • Intermediate stabilization – often electrostatic • Perfect geometry – easy to observe, difficult to recreate(Knowles 1991).
E-S encounter: a simple but important problem • Principal mechanism: Create an attractive electrostatic field near the active site, even if E and S are Coulombically repulsed • Some E-S have been designed to have super-perfect (~109/s) encounter rates (e.g. SOD/superoxide, barnase/barstar) • Precise details still being elucidated, but
To understand an enzyme: Progress and future directions Enzyme Design Precise structural details (crystal structure) Identify catalytic residues Identify enzyme Qualitative understanding Quantitative (QM) understanding)
Predictions for the future • Enzyme design / engineering – definitely possible, but can naturally high rates be attained? • Can any protein fold be an enzyme, or are some protein folds more suited than others? • Quantum level understanding – may help in optimizing enzymes, but not a critical part of basic understanding
Acknowledgements • Dr. Jim Stivers • Dr. Marc Ostermeier • Dr. Jeff Gray and Gray lab members • Practice talk attendees
References General: Schreiber, G. and Fersht, A.R. (1996). Rapid, electrostatically assisted association of proteins. Nat. Struct. Biol.3 (5), 427-431. Radzicka, A. and Wolfenden, R. (1995). A proficient enzyme. Science267 (5194), 90-93. Hiromi, K. (1979). Kinetics of Fast Enzyme Reactions. Kodansha Ltd., Tokyo. Alberty, R.A. and Hammes, G.G. (1958). Application of the theory of diffusion-controlled reactions to enzyme kinetics. J. Phys. Chem.62, 154-159. Carbonic Anhydrase: Lindskog, S. (1997). Structure and mechanism of carbonic anhydrase. Pharmacol. Ther. 74(1), 1-20. Jackman, J.E., Merz K.M. Jr., Fierke, C.A. (1996). Disruption of the active site solvent network in carbonic anhydrase II decreases the efficiency of proton transfer. Biochemistry35 (51): 16421-8. Krebs, J.F., Ippolito, J.A., Christianson, D.W., Fierke, C.A. (1993). Structural and functional importance of a conserved hydrogen bond network in human carbonic anhydrase II.
Xue, Y., Liljas, A., Jonsson, B.H., and Lindskog, S. (1993). Structural analysis of the zinc hydroxide-Thr199-Glu106 hydrogen bond network in human carbonic anhydrase II. Proteins17(1), 93-106. Tu, C., Silverman, D.N., Forsman, C., Jonsson, B.H., and Lindskog, S. (1989). Role of histidine 64 in carbonic anhydrase II studied with a Site-Specific Mutant. Biochemistry28, 7913-7918. Triose Phosphate Isomerase: Jogl, G., Rozovsky, S., McDermott, A.E., Tong, L. (2003). Optimal alignment for enzymatic proton transfer: structure of the Michaelis complex of triosephosphate isomerase at 1.2A resolution. PNAS100(1), 50-55. Kursula, I. and Wierenga, R.K. (2003). Crystal structure of triosephosphate isomerase complexed with 2-phosphoglycolate at 0.83A resolution. J. Biol. Chem. 278 (11), 9544-51. Cui, Q. and Karplus, M. (2001). Triosephosphate isomerase: a theoretical comparison of alternative pathways. JACS123 (10), 2284-90. Wade, R.C., Gabdoulline, R.R., Luty, B.A. (1998). Species dependence of enzyme-substrate encounter rates for triose phosphate isomerase. Proteins31, 406-416.
Harris, T.K., Abeygunawardana, C., Mildivan, A.S. (1997). NMR studies of the role of hydrogen bonding in the mechanism of triosephosphate isomerase. Biochemistry36 (48), 14661-75. Komives, E.A., Chang, L.C., Lolis, E., Tilton, R.F., Petsko, G.A., Knowles, J.R. Electrophilic catalysis in triosephosphate isomerase: the role of histidine 95. Biochemistry30 (12), 3011-9. Lodi, P.J. and Knowles, J.R. (1991). Neutral imidazole is the electrophile in the reaction catalyzed by triosephosphate isomerase: structural origins and catalytic applications. Biochemistry30 (28), 6948-56.