Download

1 / 46
480 likes | 763 Views
Lipids, Biological Membranes, and Membrane Transport Chapters 9 and 10. The lipid bilayer. The thickness of a bilayer is usually up to around 60 Å. Is the barrier that keeps ions, proteins and other molecules where they are needed.

E N D
Lipids, Biological Membranes, and Membrane TransportChapters 9 and 10
The lipid bilayer • The thickness of a bilayer is usually up to around 60 Å. • Is the barrier that keeps ions, proteins and other molecules where they are needed. • Are impermeable to most water-soluble (hydrophilic) molecules. • Particularly impermeable to ions, which allows cells to regulate salt concentrations and pH by pumping ions across their membranes using proteins called ion pumps.
Biological Membranes • Typically include several types of lipids other than phospholipids. • A particularly important example in animal cells is cholesterol, which helps strengthen the bilayer and decrease its permeability • When closed into `bubbles', bilayers provide a barrier between `inside' and `outside'; i.e. they define closed `compartments'. • Large bubbles (microns in diameter) are often called `vesicles'
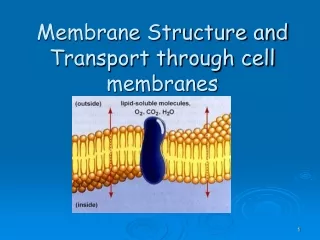
The Plasma Membrane • Composed of a phospholipid bilayer and proteins. • The phospholipid sets up the bilayer structure • Phospholipids have • hydrophilic heads and fatty acid tails. • The plasma membrane is fluid--that is proteins move in a fluid lipid background
The Fluid Mosaic Model Originally proposed by S. Jonathan Singer and Garth Nicolson in 1972. Allows for dynamic nature of membrane Little transition of lipids can take place without specific enzymes to mediate transfer - flippase.
Flippase • Enzymes located in the membrane responsible for aiding the movement of phospholipid molecules between the two leaflets that compose a cell's membrane • Two types: • Transverse • Lateral
Transverse Diffusion Or flip-flop involves the movement of a lipid or protein from one membrane surface to the other. Is a fairly slow process due to the fact that a relatively significant amount of energy is required for flip-flopping to occur.
Transverse Diffusion Most large proteins do not flip-flop due to their extensive polar regions, which are unfavorable in the hydrophobic core of a membrane bilayer. This allows the asymmetry of membranes to be retained for long periods, which is an important aspect of cell regulation.
Lateral Diffusion • Refers to the lateral movement of lipids and proteins found in the membrane. • Membrane lipids and proteins are generally free to move laterally if they are not restricted by certain interactions. • Is a fairly quick and spontaneous process
The Endo-membrane system • Proteins or lipids made in the ER contained in transport vesicles fuse with the Golgi. • The Golgi modifies proteins and lipids from the ER, sorts them and packages them into transport vesicles. • This transport vesicle “buds off” and moves to the cytoplasm. • Fuse with plasma membrane.
Flippase • Potential role of ATP-dependent lipid flippases in vesicle formation. • ATP-dependent lipid translocation might help deform the membrane by moving lipid mass towards the cytoplasmic leaflet
Flippase • This area asymmetry will increase the spontaneous curvature of the bilayer, and may thus help deform the membrane during vesicle budding. • Lem3-Cdc50 proteins regulate the localization and activity of P4-ATPases. • P4-ATPases play a pivotal role in the biogenesis of intracellular transport vesicles, polarized protein transport and protein maturation.
Flippase • Interaction of P4-ATPases with peripheral guanine nucleotide-exchange factors (GEFs) might cause activation of small GTPases. • GTPases subsequently bind to the membrane and facilitate the assembly of coat proteins (if required) • And thus, the endo-membrane system allows gene expression, post-translational modification, and secretion to occur!
Membrane Structure and Dynamics Membrane functions - physical barrier from entry and exit form cell and organelles
Phospholipids: Two fatty acids covalently linked to a glycerol, which is linked to a phosphate. All attached to a “head group”, such as choline, an amino acid. Head group POLAR – so hydrophilic (loves water) Tail is non-polar –hydrophobic The tail varies in length from 14 to 28 carbons.
Membrane components - • 60 to 70% of mammalian lipids are phospholipids • Bacteria have almost no PC and are mostly PE • Neuronal tissue (myelin) PI > PC • Alterations in lipid composition - permeability, fluidity, exocytosis, neural transmission and signaling potential
Membrane Asymmetry • P-ethanolamine and P-serine predominately faces inside of cell • P-choline faces outside of membrane and inside of organelles • carbohydrates of glycoproteins facing outside • During apoptosis there is a re-arraignment of lipids where phosphatidyl serine moves to the exterior face of the membrane. • One of the key signals of cell death
Proteins - Add function and structure to membrane • Extrinsic proteins (peripheral) • Loosely attached to membrane • ionic bonds with polar head groups and carbohydrates • hydrophobic bonds with lipid • proteins have lipids tails • easily displaced from membrane • salt, pH, sonication
Integral proteins - tightly bound to membrane - span both sides Protein has both polar and hydrophobic sections removed only through disrupting membrane with detergents detergents disrupt lipid bilayer and incorporate proteins and some lipids into detergent micelles • allows for purification of membrane proteins • reconstitute into specific vesicles for study
Transmembrane proteins • So designated because they are both structurally and functionally an integral component of a membrane. • Example: • Human erythrocyte glycophorin A • Involved in interactions with the Red Blood Cell cytoskeleton that may modulate membrane rigidity. • The extracellular portion of the protein also serves as the receptor for the influenza virus
Transmembrane proteins • Has a total molecular weight of about 31,000 and is approximately 40% protein and 60% carbohydrate. • The primary structure consists of a segment of 19 hydrophobic amino acid residues with a short hydrophilic sequence on one end and a longer hydrophilic sequence on the other end. • The 19-residue sequence is just the right length to span the cell membrane if it is coiled in the shape of an α-helix. • The large hydrophilic sequence includes the amino terminal residue of the polypeptide chain.
Transmembrane proteins • General “Rules of thumb” • takes about 20 aa to cross membrane • many proteins cross many times • odd # of transmembrane regions, • -COOH terminal usually cytosolic • -NH3+ terminal extracellular • can be predicted by amino acid sequence • high % of side chains will be hydrophobic
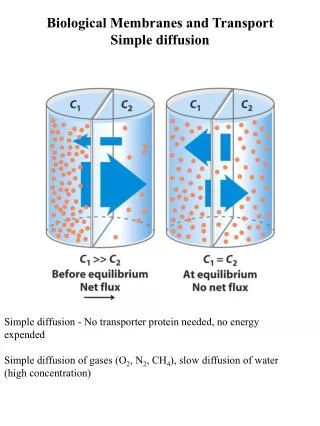
membrane transport • The term refers to the collection of mechanisms that regulate the passage of solutes such as ions and small molecules through biological membranes namely lipid bilayers that contain proteins embedded in them. • The regulation of passage through the membrane is due to selective membrane permeability. • The movements of most solutes through the membrane are mediated by membrane transport proteins which are specialized to varying degrees in the transport of specific molecules.
An example of membrane transport • Cholesterol • waxy steroid metabolite found in the cell membranes and transported in the blood plasma of all animals. • Essential structural component of mammalian cell membranes, required to establish proper membrane permeability and fluidity • transported in the circulatory system within lipoproteins • LDL molecules are the major carriers of cholesterol in the blood • each one contains approximately 1,500 molecules of cholesterol • Recognized by the LDL receptor
An example of membrane transport • Upon binding many LDL receptors become localized in clathrin-coated pits. • Both the LDL and its receptor are internalized by endocytosis to form a vesicle within the cell. • The vesicle then fuses with a lysosome, which has an enzyme called lysosomal acid lipase that hydrolyzes the cholesterol. • Now within the cell, the cholesterol can be used for membrane biosynthesis or esterified and stored within the cell, so as to not interfere with cell membranes.
Cholesterol Uptake • Cells destined to take up cholesterol possess surface receptors for the LDL particle. • Receptor Binding & Activation: that LDL receptor binds to Apo-B protein on the LDL particle • Coated Pit Formation: • Clathrin forms cage around forming endosome • Clathrin-Coated Vesicle Budding • Uncoating of the Vesicle
Cholesterol Uptake • Early Endosome associates with other vesicle • Formation of CURL (Compartment for Uncoupling of Ligand and Receptor) or Late Endosome • Recycling of the Receptor to the cell surface • Fusion of Transport Vesicle with Lysosome • Digestion of the LDL to Release Cholesterol
CURL Formation & Lysosome Digestion • CURL = Compartment for Uncoupling of Receptor & Ligand • pH drops to acidic (pH 5) • Conformational change in Receptor releases LDL • Receptor recycles to cell membrane • Late Endosome fuses with lysosomal vesicles • LDL is degraded; Esterases digest esters Cholesterol is released into the cytoplasm
Familial Hypercholesterolemia (FH): high levels of blood cholesterol and other characteristics • Leads to an increase in blood LDL (cholesterol) • Risk of Atherosclerosis & Heart Disease • Atherosclerosis: buildup of cholesterol deposits lead to plaques & clog arteries • Contributes to heart attacks at early age • One human mutation is due to a defect in LDL receptor (e.g., in adapter binding site; can't form coated pit for LDL uptake) which causes the buildup of LDL particles at the cell surface leading to plaque formation.
Summary of membrane transport • Three types of membrane transporters enhance the movement of solutes across plant cell membranes • Channels – passive transport • Carriers – passive transport • Pumps- active transport
Channels • Transmembrane proteins that work as selective pores • Transport through these passive • The size of the pore determines its transport specifity • Movement down the gradient in electrochemical potential • Unidirectional • Very fast transport • Limited to ions and water
Channels • Sometimes channel transport involves transient binding of the solute to the channel protein • Channel proteins have structures called gates. • Open and close pore in response to signals • Light • Hormone binding • Only potassium can diffuse either inward or outward • All others must be expelled by active transport.
The aquaporin channel protein • There is some diffusion of water directly across the bi-lipid membrane. • Aquaporins: Integral membrane proteins that form water selective channels – allows water to diffuse faster • Facilitates water movement in plants • Alters the rate of water flow across the plant cell membrane – NOT direction
Carriers • Do not have pores that extend completely across membrane • Substance being transported is initially bound to a specific site on the carrier protein • Carriers are specialized to carry a specific organic compound • Binding of a molecule causes the carrier protein to change shape • This exposes the molecule to the solution on the other side of the membrane • Transport complete after dissociation of molecule and carrier protein
Carriers • Moderate speed • Slower than in a channel • Binding to carrier protein is like enzyme binding site action • Can be either active or passive • Passive action is sometimes called facilitated diffusion • Unidirectional
Example: GLUT1 glucose carrier • GLUT1 is a large integral protein, predicted via hydropathy plots to include 12 transmembrane a-helices • Transporter exists in 2 conformations, T1 with glucose binding site exposed on outer surface of plasma membrane, and T2, with binding site exposed on inner surface. • D-Glc binding on T1 triggers change to T2. • Glc is released into cytosol, triggering conformational change back to T1, ready to pick up another glucose from the outside. • Process is fully reversible, and as [S]in approaches [S]out, rates of entry and exit become equal.
Active transport • To carry out active transport: • The membrane transporter must couple the uphill transport of a molecule with an energy releasing event • This is called Primary active transport • Energy source can be • The electron transport chain of mitochondria • The electron transport chain of chloroplasts • Absorption of light by the membrane transporter • Such membrane transporters are called PUMPS
Primary active transport- Pumps • Movement against the electrochemical gradient • Unidirectional • Very slow • Significant interaction with solute • Direct energy expenditure
pump-mediated transport against the gradient (secondary active transport) • Involves the coupling of the uphill transport of a molecule with the downhill transport of another • (A) the initial conformation allows a proton from outside to bind to pump protein • (B) Proton binding alters the shape of the protein to allow the molecule [S] to bind
pump-mediated transport against the gradient (secondary active transport) • (C) The binding of the molecule [S] again alters the shape of the pump protein. This exposes the both binding sites, and the proton and molecule [S] to the inside of the cell • (D) This release restores both pump proteins to their original conformation and the cycle begins again
pump-mediated transport against the gradient (secondary active transport) • Two types: • (A) Symport: • Both substances move in the same direction across membrane • (B) Antiport: • Coupled transport in which the downhill movement of a proton drives the active (uphill) movement of a molecule
pump-mediated transport against the gradient (secondary active transport) • The proton gradient required for secondary active transport is provided by the activity of the electrogenic pumps • Membrane potential contributes to secondary active transport • Passive transport with respect to H+ (proton)