Download
1 / 1
10 likes | 158 Views
Voelckel C*, Biggs P, Deusch O, Lockhart PJ *Allan Wilson Centre for Molecular Ecology and Evolution, Massey University, Palmerston North, New Zealand c.voelckel@massey.ac.nz.

E N D
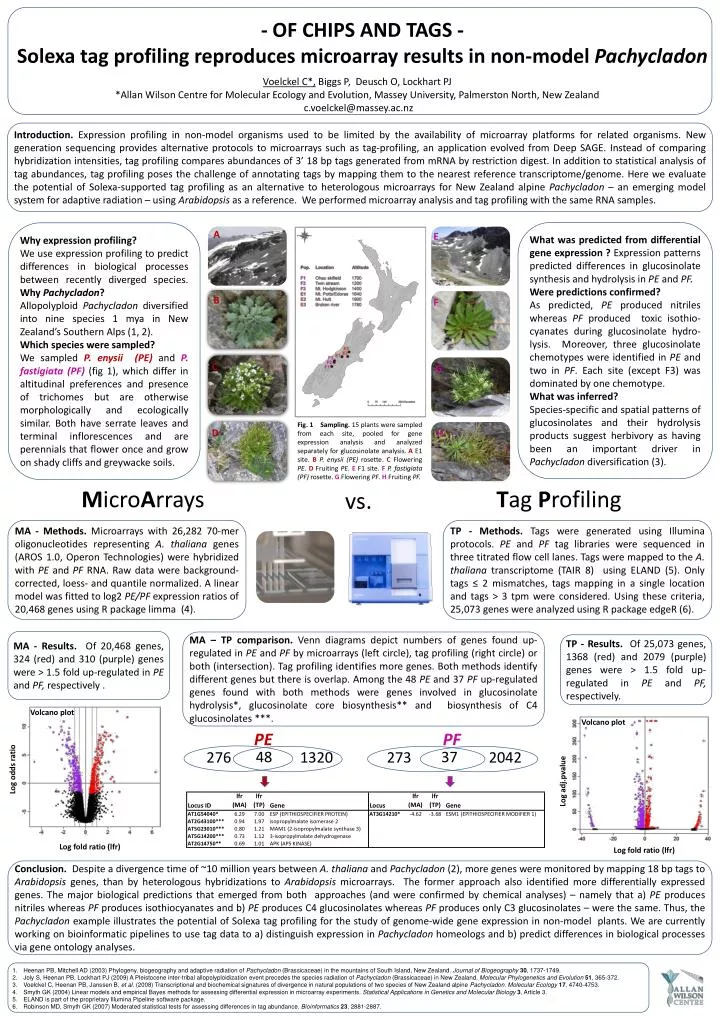
Voelckel C*, Biggs P, Deusch O, Lockhart PJ *Allan Wilson Centre for Molecular Ecology and Evolution, Massey University, Palmerston North, New Zealand c.voelckel@massey.ac.nz Introduction. Expression profiling in non-model organisms used to be limited by the availability of microarray platforms for related organisms. New generation sequencing provides alternative protocols to microarrays such as tag-profiling, an application evolved from Deep SAGE. Instead of comparing hybridization intensities, tag profiling compares abundances of 3’ 18 bp tags generated from mRNA by restriction digest. In addition to statistical analysis of tag abundances, tag profiling poses the challenge of annotating tags by mapping them to the nearest reference transcriptome/genome. Here we evaluate the potential of Solexa-supported tag profiling as an alternative to heterologous microarrays for New Zealand alpine Pachycladon – an emerging model system for adaptive radiation– using Arabidopsis as a reference. We performed microarray analysis and tag profiling with the same RNA samples. What was predicted from differential gene expression ? Expression patterns predicted differences in glucosinolate synthesis and hydrolysis in PE and PF. Were predictions confirmed? As predicted, PE produced nitriles whereas PF produced toxic isothio-cyanates during glucosinolate hydro-lysis. Moreover, three glucosinolatechemotypes were identified in PE and two in PF. Each site (except F3) was dominated by one chemotype. What was inferred? Species-specific and spatial patterns of glucosinolates and their hydrolysis products suggest herbivory as having been an important driver in Pachycladon diversification (3). Why expression profiling? We use expression profiling to predict differences in biological processes between recently diverged species. Why Pachycladon? Allopolyploid Pachycladon diversified into nine species 1 mya in New Zealand’s Southern Alps (1, 2). Which species were sampled? We sampled P. enysii (PE) and P. fastigiata (PF)(fig 1), which differ in altitudinal preferences and presence of trichomes but are otherwise morphologically and ecologically similar. Both have serrate leaves and terminal inflorescences and are perennials that flower once and grow on shady cliffs and greywacke soils. A E B F - OF CHIPS AND TAGS - Solexa tag profiling reproduces microarray results in non-model Pachycladon C G Fig. 1 Sampling. 15 plants were sampled from each site, pooled for gene expression analysis and analyzed separately for glucosinolate analysis. A E1 site. BP. enysii (PE) rosette. CFlowering PE. D Fruiting PE. E F1 site. FP. fastigiata (PF) rosette. G Flowering PF. H Fruiting PF. D H MicroArrays Tag Profiling vs. MA - Methods. Microarrays with 26,282 70-mer oligonucleotides representing A. thalianagenes (AROS 1.0, OperonTechnologies) were hybridized with PE and PFRNA. Raw data were background-corrected, loess- and quantile normalized. A linear model was fitted to log2 PE/PF expression ratios of 20,468 genes using R package limma (4). TP - Methods. Tags were generated using Illumina protocols. PE and PF tag libraries were sequenced in three titrated flow cell lanes. Tags were mapped to the A. thalianatranscriptome (TAIR 8) using ELAND (5). Only tags ≤ 2 mismatches, tags mapping in a single location and tags > 3 tpm were considered. Using these criteria, 25,073 genes were analyzed using R package edgeR (6). MA - Results. Of 20,468 genes, 324 (red) and 310 (purple) genes were > 1.5 fold up-regulated in PE and PF, respectively . MA – TP comparison. Venn diagrams depict numbers of genes found up-regulated in PE and PF by microarrays (left circle), tag profiling (right circle) or both (intersection). Tag profiling identifies more genes. Both methods identify different genes but there is overlap. Among the 48 PE and 37 PF up-regulated genes found with both methods were genes involved in glucosinolate hydrolysis*, glucosinolate core biosynthesis** and biosynthesis of C4 glucosinolates ***. TP - Results. Of 25,073 genes, 1368 (red) and 2079 (purple) genes were > 1.5 fold up-regulated in PE and PF, respectively. Volcano plot Volcano plot PE PF 48 37 276 1320 273 2042 Log odds ratio Log adj.pvalue Log fold ratio (lfr) Log fold ratio (lfr) Conclusion. Despite a divergence time of ~10 million years between A. thaliana and Pachycladon(2), more genes were monitored by mapping 18 bp tags to Arabidopsis genes, than by heterologoushybridizations to Arabidopsis microarrays. The former approach also identified more differentially expressed genes. The major biological predictions that emerged from both approaches (and were confirmed by chemical analyses) – namely that a) PE produces nitriles whereas PF produces isothiocyanates and b) PEproduces C4 glucosinolates whereas PF produces only C3 glucosinolates – were the same. Thus, the Pachycladon example illustrates the potential of Solexa tag profiling for the study of genome-wide gene expression in non-model plants. We are currently working on bioinformatic pipelines to use tag data to a) distinguish expression in Pachycladonhomeologs and b) predict differences in biological processes via gene ontology analyses. Heenan PB, Mitchell AD (2003) Phylogeny, biogeography and adaptive radiation of Pachycladon(Brassicaceae) in the mountains of South Island, New Zealand. Journal of Biogeography30, 1737-1749. Joly S, Heenan PB, Lockhart PJ (2009) A Pleistocene inter-tribal allopolyploidization event precedes the species radiation of Pachycladon(Brassicaceae) in New Zealand. Molecular Phylogenetics and Evolution51, 365-372. Voelckel C, Heenan PB, Janssen B, et al. (2008) Transcriptional and biochemical signatures of divergence in natural populations of two species of New Zealand alpine Pachycladon. Molecular Ecology17, 4740-4753. Smyth GK (2004) Linear models and empirical Bayes methods for assessing differential expression in microarray experiments. Statistical Applications in Genetics and Molecular Biology3, Article 3. ELAND is part of the proprietary Illumina Pipeline software package. Robinson MD, Smyth GK (2007) Moderated statistical tests for assessing differences in tag abundance. Bioinformatics 23, 2881-2887.